Ministry of Education Key Laboratory for Ecology of Tropical Islands, Key Laboratory of Tropical Animal and Plant Ecology of Hainan Province, College of Life Sciences, Hainan Normal University, Haikou, 571158, China
Funds: This work was supported by the National Natural Science Foundation of China (No. 31970427 and 32270526 to WL). WL was supported by the specific research fund of The Innovation Platform for Academicians of Hainan Province
Most studies exploring abilities of hosts to detect brood parasitism are based on detecting colour and/or pattern differences among parasitic and host eggs or nestlings, while only few were focused on size differences. True recognition and recognition by discordancy are used to explain cognitive mechanisms of host egg recognition; however, only a few studies have found that hosts use recognition by discordancy. This study investigated: 1) whether egg and nestling recognitions in the Crested Myna (Acridotheres cristatellus) are based on size cues; 2) whether the egg cognitive mechanism is recognition by discordancy based on size cues; and 3) whether the longer the experiment time, the higher the egg recognition rate. Our results showed that the Crested Myna uses egg or nestling size as a recognition cue while the egg and nestling colour and patterning are not associated with egg or nestling rejection, thus the cognitive mechanism of egg recognition in the Crested Myna is recognition by discordancy based on egg size cues. Furthermore, there is a rejection delay in time of egg rejection behaviour of the Crested Myna. Therefore, we suggest that the periodicity of egg rejection experiments could be appropriately extended, especially for species with relatively low egg recognition ability.
The host ability of recognizing and rejecting parasitic eggs has been the focus of most previous studies on parasite–host systems. Host egg rejection is mainly achieved through the ejection of eggs (Sealy and Neudorf, 1995; Zhang et al., 2021), burial of eggs (Moskát and Honza, 2002; Guigueno and Sealy, 2010), or nest desertion (Goguen and Mathews, 1996; Hosoi and Rothstein, 2000). However, different hosts vary in their ability to recognise eggs (Davies and Brooke, 1989; Davies, 2000), and one possible reason for this variation is that hosts differ in their ability to perceive morphological contrast between parasitic eggs and their own eggs (Kilner, 2006). Parasitic eggs produce multimodal and multicomponent recognition cues, and both tactile (size, shape, and texture) and visual (size, shape, colour, and spots) patterns can be involved in host perception and recognition of foreign eggs (Moskát et al., 2008; Honza and Cherry, 2017; Hanley et al., 2019). The relative contributions of egg size and eggshell shape (e.g., colour and spottiness) to host egg recognition and rejection show substantial variation across parasite–host species and systems (Segura et al., 2016; Honza and Cherry, 2017; Li et al., 2020). Although the role of eggshell shape in parasitic egg recognition by hosts has received some support (Krüger, 2007; Polačikova et al., 2007; Holveck et al., 2010; Honza and Cherry, 2017; Abolins-Abols et al., 2019), the role of egg size has been more scarcely studied, mainly in host of cowbirds (Ye et al., 2022). In cuckoo–host systems, most studies have shown that egg size and egg colour/pattern have mixed effects on egg recognition, while the importance of egg size recognition as a specific antiparasitic adaptation is unclear.
Egg rejection is a complex decision-making process in which the host responds to cognitive stimuli from alien eggs (Antonov et al., 2008; Soler et al., 2017), consisting of three stages: egg recognition, the decision of whether or not to reject the putative parasitic egg and the act of ejection itself (Soler et al., 2012). Theoretical models and empirical data suggest that there is a threshold in host cognitive abilities that minimise the risk of recognition errors (Lotem et al., 1995; Davies et al., 1996; Antonov et al., 2008). Most hosts know the appearance of their eggs and reject foreign eggs (true recognition); a few hosts recognise foreign eggs based on size or shape differences (recognition by discordancy), and it is easy to misidentify the alien eggs (Yi, 2020). The delay in rejection of foreign eggs might indicate perceptual problems influencing decision making and, thus, level of antiparasitic defences of individual hosts (Antonov et al., 2008; Požgayová et al., 2011). Some individuals may eject a foreign egg the same day it is introduced into the nest (even immediately, within minutes or even seconds), while others delay their response for several days (Sealy and Neudorf, 1995; Honza et al., 2005; Antonov et al., 2008; Požgayová et al., 2011; Soler et al., 2017). As far as we know, the delay of egg rejection has received little attention.
The behavioural mechanisms of nest sanitation (the removal of nestling faeces, eggshells, unhatched eggs, dead nestlings, and foreign material from the nest) is very similar to egg ejection (Guigueno and Sealy, 2009; 2012; Luro and Hauber, 2017). Rothstein (1975) first suggested that nest-clearing behaviour in birds could be a pre-adaptation facilitating the evolution of egg rejection. Based on this hypothesis, Moskát et al. (2003) proposed a hierarchical process to explain the connection between nest sanitation and egg-rejection behaviour as follows: 1) hosts reject all non-egg-shaped objects as a general cleaning mechanism; 2) hosts with egg recognition ability can distinguish between egg analogues and parasitic eggs. In other words, the less the foreign object in the nest resembles an egg, the more discriminative the bird is, and the higher the likelihood that the foreign object will be rejected. This prediction is supported by some (Guigueno and Sealy, 2012; Yang et al., 2015a; 2015b; Feng et al., 2019), but not all studies (Peer and Sealy, 2004; Luro and Hauber, 2017; Su et al., 2018; Peer, 2017; Li et al., 2021), and, thus, the association between nest sanitation and egg rejection abilities is unclear at best (Luro and Hauber, 2017). Therefore, more research is needed to explore the association between nest sanitation and egg rejection.
Host birds have strong recognition abilities at the egg stage, yet curiously, most hosts do not recognise parasitic nestlings that differ markedly in appearance from their own at the nestling stage (Grim, 2006; Soler, 2014; Yang et al., 2015c), but, certain host birds do (Langmore et al., 2003; Sato et al., 2010; Tokue and Ueda, 2010; Yang et al., 2015c; Noh et al., 2018). Thus, in addition to egg recognition as an antiparasitic means of limiting cuckoo parasitism in most hosts, nestling recognition is also an important factor limiting cuckoo parasitism, but nestling recognition in the host has received less attention.
We conducted four types of experiments between 2018 and 2022 with the Crested Myna (Acridotheres cristatellus) as follows: (1) nest sanitation experiments, in which non-oval peanuts or oval cherry tomatoes were added to the nest; we predict a high rejection rate for peanuts. (2) Egg rejection experiments, in which red, white, and white spotted Crested Myna model eggs or real eggs of different sizes (large, medium, and small) were added to the nest; we predict no significant difference in rejection rates of red, white, and white spotted Crested Myna model eggs, but the rejection rate of small eggs should be significantly higher, which supports the parental investment theory. (3) Egg recognition mechanism experiments, the number of different types of eggs were manipulated, in which “3 + 1” and “2 + 2” exchange experiments were performed; we predict that if Crested Mynas are using a discordancy recognition mechanism they should reject the egg types that are less abundant in the nest. (4) Nestling recognition experiments, in which the Crested Myna nestlings of different sizes (large and small) or differently sized (large and small) heterospecific nestlings were added to the nest; we predict that mynas rejects small size heterospecific nestlings. In addition, we investigated the effect of different experimental periods (long and short time) on egg rejection, and predicted high egg rejection rates for long experimental periods.
2.
Methods
2.1
Study area and study species
The study was conducted in Ding’an County (19°28′ N, 110°24′ E) and Tanniu Town, Wenchang City (19°40′ N, 110°45′ E), Hainan Island, southern China. The two sites are approximately 40 km apart, and both are relatively closed agroecosystems. Both sites have a tropical monsoonal maritime climate with abundant sunshine and rainfall (http://dingan.hainan.gov.cn/dingan/zjda/dagk/201111/t20111121_1681790.html; http://wenchang.hainan.gov.cn/wenchang/dlzy/list_tt.shtml). Tree holes in both locations are a restricted and scarce resource, as trees that become old or decayed are often cut down in time by local farmers. The availability of artificial nest boxes can be effective in alleviating the limitations of nest site availability for birds, and the birds that breed using artificial nest boxes at both sites mainly include the Crested Myna, Common Myna (Acridotheres tristis), Oriental Magpie Robin (Copsychus saularis), and White-shouldered Starling (Sturnus sinensis).
The Crested Myna is a secondary cavity-nesting bird that is widely distributed in south-central and south-eastern China, as well as in India and Vietnam (Craig and Feare, 1998; Zheng, 2017). The Hainan subspecies of the Crested Myna (subspecies Acridotheres c. brevipennis) is a common resident bird on Hainan Island (Zheng, 2017) and can breed on high coconut treetops or with the aid of artificial nest boxes (Fig. 1; see also Liu and Liang, 2021). The Crested Myna lays one egg per day, which is pure blue-green in colour, with a clutch size of mostly four eggs and an incubation period of approximately 12 days (breeding information see Liu and Liang, 2021). To the best of our knowledge, few studies on Crested Mynas (about 23 studies, data from Web of Science by searching “Acridotheres cristatellus”) have focused on bird community structure, dispersal, or physiology. We regard Crested Mynas as the potential host of cuckoos, although there is no direct evidence, because: 1) the breeding information of Crested Mynas is limited, and breeding information in natural caves has not been studied; 2) Common Mynas (nests, eggs or incubation and brooding behaviours are similar to Crested Mynas) are the host of Asian Koels (Eudynamys scolopaceus) and Great Spotted Cuckoos (Clamator glandarius), and the size is used as a cue for egg recognition (Sadam, 2022).
Figure
1.
Natural nest of the Crested Myna (A); nest of the Crested Myna in an artificial nest box (B); one peanut added to the Crested Myna nest (C); one cherry tomato added to the Crested Myna nest (D).
In this study, field data collection was conducted during the breeding season of the Crested Myna (from March to August). Wooden nest boxes were hung at a height of approximately 3 m above the ground in early March each year. Small nest boxes (L × W × H = 15 cm × 15 cm × 30 cm) were hung from 2018 to 2022, large nest boxes (L × W × H = 25 cm × 25 cm × 30 cm) were hung in 2022. For egg rejection experiments during the early incubation stage of the Crested Myna (day ≤ 3), we added one experimental egg to the nest and did not remove the Crested Myna eggs from the nest, as several studies have shown that this is a normal procedure for experiments and does not alter host rejection rates (Davies and de Brooke, 1989; Grim and Honza, 2001; Li et al., 2020). The experimental period for egg rejection experiments is typically 6 days (Luro et al., 2018; Li et al., 2020; Ye et al., 2022), but given the phenomenon of delayed egg rejection, we chose to end the experiment when the Crested Myna eggs hatched. Hosts were considered to have rejected experimental eggs if they were removed or buried or if they had visible peck marks during the experiment, and conversely, they were considered to have accepted the experimental eggs if they remained intact in the active nest (Ye et al., 2022). Similarly, we conducted experiments on egg recognition mechanisms during the early incubation stage (day ≤ 3) of the Crested Myna and ended the experiment when the Crested Myna eggs hatched. The Crested Myna eggs removed during the experiment were tagged and placed in an incubator for incubation and then placed in the corresponding nest to be fed by the Crested Myna parents after the eggs hatched. For the nestling recognition experiment, we chose to include nestlings of Grey-backed Starlings or Oriental Magpie Robins that differed significantly in appearance from the Crested Myna nestlings. The experimental period was 10 days, covering half of the nestling period of Crested Myna (20 days; Liu and Liang, 2021), and hosts were considered to have rejected the alien nestlings if they died or disappeared during the experimental period. Alien nestlings that remained alive in the nest at the end of the experiment were released back to their original nest sites.
2.3
Artificial parasitism experiments
We used polymer clay (Ai Tao Le, Shenzhen, China) of different colours (red and white) to produce the Crested Myna model eggs (28.74 ± 1.20 mm × 21.13 ± 0.53 mm; Liu and Liang, 2021), egg spots are drawn with a black marker (zebra, Shanghai, China). For comparison with the Crested Myna eggs, we chose real eggs of three different sizes for egg rejection experiments, namely Japanese Quail eggs (Coturnix japonica; large eggs), Cockatiel eggs (Nymphicus hollandicus; medium eggs), and Budgie eggs (Melopsittacus undulatus; small eggs) (Table 1). First, we explored the effect of the experimental period on Crested Myna egg rejection by adding one Budgie egg to the nest for 6 days (n = 26, small boxes) or until the Crested Myna eggs hatched (n = 24, small and large boxes). We then conducted four types of experiments as follows. (1) For nest sanitation experiments, we added one non-oval peanut (n = 19) or ovoid cherry tomato (n = 13; Fig. 1) to the small nest in 2022 during the early incubation stage (day ≤ 3) of the Crested Myna and set the experimental period to 6 days owing to the short period of preservation. (2) For egg rejection experiments, to test the effects of egg colour and egg spots on the egg rejection behaviour of the Crested Myna, a Crested Myna egg-sized red model egg (RM, n = 14), white model egg (WM, n = 12), or white spotted model egg (WSM, n = 16; Fig. 2) was added to the small nest in 2021. To test the effect of egg size on the egg rejection behaviour of the Crested Myna, a Japanese Quail egg (large egg, n = 18, abbreviated LE), Cockatiel egg (medium egg, n = 17, abbreviated ME), or Budgie egg (small egg, n = 14, abbreviated SE; Fig. 2) was added to the small nest in 2022. (3) For egg recognition mechanism experiments, to test whether egg size cues are a recognition mechanism for the Crested Myna, we performed four sets of egg-swapping experiments in 2022 as follows: a) replacing Crested Myna eggs with three Japanese Quail eggs (large) + one Crested Myna egg (small, n = 16); b) replacing Crested Myna eggs with three Japanese Quail eggs (large) + one Cockatiel egg (small, n = 15); c) replacing Crested Myna eggs with three Cockatiel eggs (small) + one Crested Myna egg (large, n = 21); and d) replacing Crested Myna eggs with two Japanese Quail eggs (large) + two Cockatiel eggs (small, n = 14) (Fig. 2). (4) For nestling recognition experiments, to test whether the Crested Myna can recognise nestlings based on cues of nestling size, we performed heterospecific and conspecific addition experiments in 2022 as follows: e) addition of one Grey-backed Starling or Oriental Magpie Robin (9 days, n = 3 or n = 3, large) to the Crested Myna (3 days, small) nest; f) addition of one Oriental Magpie Robin (3 days, n = 2, small) to the Crested Myna (3 days, large) nest or one Oriental Magpie Robin (6 days, n = 2, small) to the Crested Myna (6 days, large) nest; g) addition of one Crested Myna (3 days, n = 4, small) to the Crested Myna (6 days, large) nest; h) addition of one Crested Myna (6 days, n = 4, large) to the Crested Myna (3 days, small) nest (Fig. 3).
Table
1.
Analysis of variance comparing egg parameters among the four real eggs, using the Bonferroni method for pairwise comparisons.
Figure
2.
Crested Myna nests with Crested Myna egg-sized red, white, and white and spotted model eggs (A–C); Crested Myna nests with Japanese Quail eggs, Cockatiel eggs, and Budgie eggs (D–F); Crested Myna nests replaced with three Japanese Quail eggs + one Crested Myna egg, three Japanese Quail eggs + one Cockatiel egg, three Cockatiel eggs + one Crested Myna egg, and two Japanese Quail eggs + two Cockatiel eggs (G–J), respectively. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
Figure
3.
Experimental operation of nestling recognition. Add one small Oriental Magpie Robin nestling (A), small Crested Myna nestling (B), large Oriental Magpie Robin nestling (C), large White-shouldered Starling nestling (D).
Fisher tests were used to analyse and compare differences in the rejection rates of oval and non-oval foreign bodies by the Crested Myna and differences in rejection rates of eggs of different colours and different spots. Fisher test was used to analyse the response of the Crested Myna to egg size. GLMM was used to analyse the responses of the Crested Myna at different experimental periods. In this model, the response of the Crested Myna (rejection or acceptance) was the response variable, the location (Wenchang or Ding’an), nest box size (large or small), clutch size, and experimental period (long or short) were fixed factors, and the nest ID was random factors. Pairwise comparisons were conducted by the least significant differences (LSD) test. For the egg recognition mechanism experiments, we treated the four groups as a whole, and Chi-square tests were used to analyse and compare the difference in the proportion of small and large eggs discarded by the Crested Myna. All tests were two-tailed, with a significance level of P < 0.05. Data are presented as the mean ± standard deviation (SD). All data analyses were performed using IBM SPSS 22.0 software (IBM Corp., Armonk, NY, USA).
3.
Results
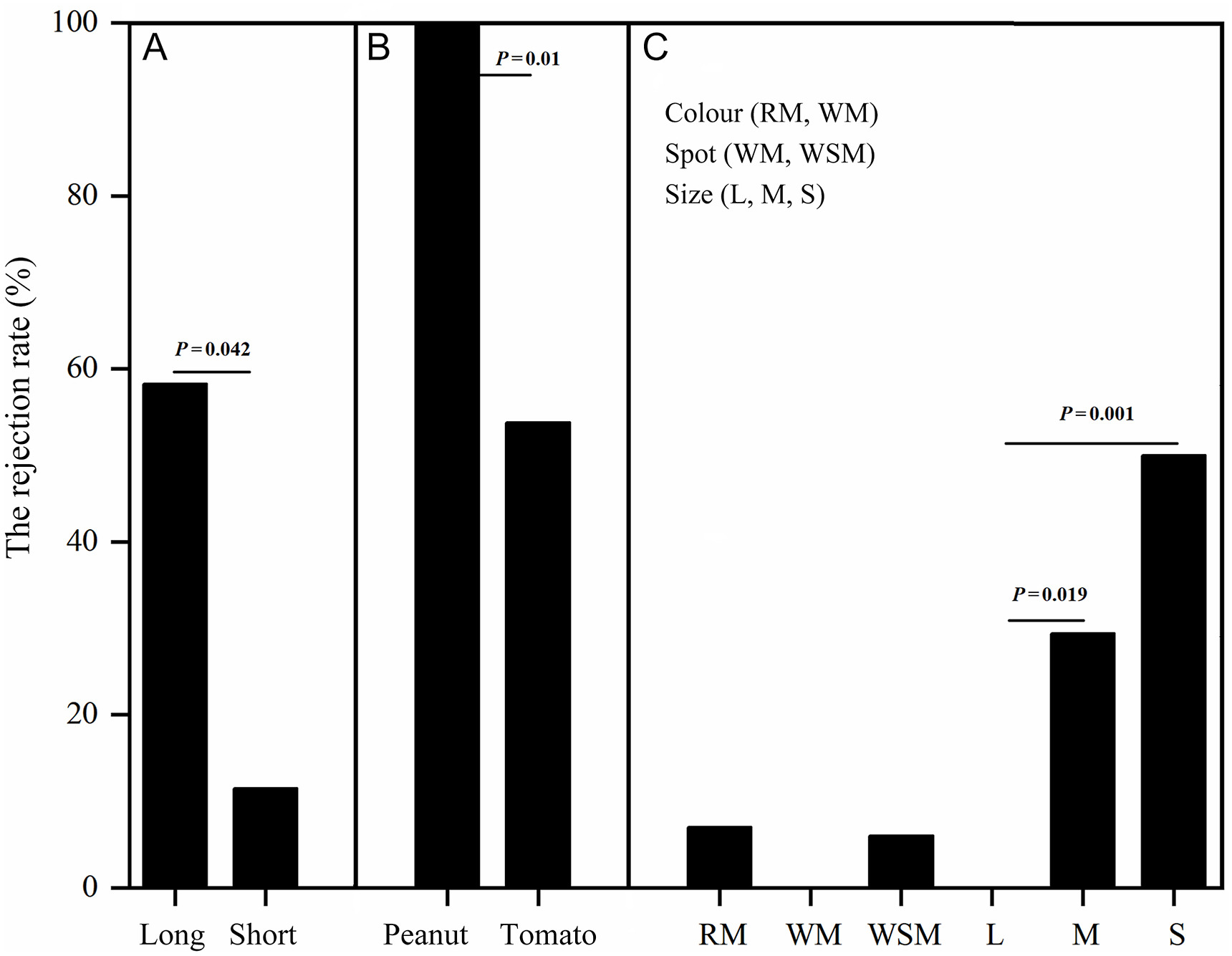
The rejection rates of the Crested Myna at different experimental periods was different (F = 3.456, P = 0.026; GLMM; Table 2), and the detected rejection rate during the short experimental period (11.5%) was lower than during the long experimental period (58.3%; P = 0.042, LSD test; Fig. 4A). For the Crested Myna, the rejection rates of peanuts (100%) was significantly higher than that with cherry tomatoes (53.8%; P = 0.01, Fisher test; Fig. 4B). There was no significant difference in the rejection rate between the RM (7.1%) and WM (0%) (P = 1, Fisher test). There was also no significant difference in rejection rates between WM (0%) and WSM (6.3%) (P = 1, Fisher test). Moreover, there was a difference in the rejection rate based on egg size (P = 0.001, Fisher test), and the rejection rates of the LE (0%) was significantly lower than that the SE (50%; P = 0.001) and the ME (29.4%; P = 0.019) (Fig. 4C). In the egg recognition mechanism experiments, the Crested Myna rejected small eggs (34.8%) more frequently than they did with large eggs (9.1%) (χ2 = 12.771, df = 1, P = 0.001, Chi-square tests; Table 3). In the nestling recognition experiments, the rejection rate was 16.7% for large-bodied heterospecific nestlings and 100% for small-bodied heterospecific nestlings; all large-bodied conspecific nestlings and small-bodied conspecific nestlings were accepted (Table 3).
Table
2.
Results of the generalised linear mixed model (GLMM) analysis of the response of the Crested Myna to different experimental time periods.
Table
3.
Results of experiments on egg recognition mechanisms and parent-nestling recognition in the Crested Myna (L refers to large and S to small size groups).
Experiment operation
All accepted
S rejected
L rejected
S and L rejected
Three quail eggs (L) + one myna (S)
7
4
4
1
Three quail eggs (L) + one cockatiel (S)
4
6
1
4
Three cockatiel eggs (S) + one myna (L)
13
6
0
2
Two quail eggs (L) + two cockatiel (S)
5
7
1
1
Myna nestlings (S) + one starling (L)
3
0
0
0
Myna nestlings (S) + one magpie robin (L)
2
0
1
0
Myna nestlings (L) + one magpie robin (S)
0
4
0
0
Myna nestlings (S) + one myna (L)
4
0
0
0
Myna nestlings (L) + one myna (S)
4
0
0
0
The number in the table refers to the number of nests.
Our experimental results showed that the Crested Myna rejected non-oval foreign bodies (peanuts) significantly more often than oval foreign bodies (cherry tomatoes), which supports the hierarchical correlation between nest sanitation and egg rejection behaviours proposed by Moskát et al. (2003). Egg rejection rates for short experimental periods were significantly lower than those for long periods; specifically, egg rejection rates increased with an increasing nest age, suggesting a temporal delay in egg rejection behaviour in the Crested Myna. Egg rejection rates were not significantly different between RM and WM or between WM and WSM, suggesting that egg colour and egg spots do not affect the egg rejection behaviour in the Crested Myna. The rejection rate with the LE was significantly lower than the rejection rates with the SE, suggesting that the Crested Myna uses egg size as a cue for recognition. In the egg recognition mechanism experiments, the percentage of small eggs in the nest rejected by the Crested Myna was significantly higher than the percentage of large eggs rejected by the Crested Myna, even for its own eggs in the small egg position, suggesting discordant recognition in this species with egg size as a recognition cue. In the nestling recognition experiments, the Crested Myna accepted conspecific nestlings of different body sizes and large heterospecific nestlings that differed greatly in appearance, but rejected small heterospecific nestlings, suggesting that Crested Mynas are likely to use nestling size as a cue for recognition.
Liu et al. (2021) found that the Red-whiskered Bulbul (Pycnonotus jocosus) is more receptive to parasitic eggs as the nest age increases. However, our results showed that the Crested Myna rejected more alien eggs as the nest age increased. The delay in egg rejection might be related to proximate mechanisms of egg recognition and could be due to problems in perceiving alien eggs during the decision-making process (Rodríguez-Gironés and Lotem, 1999; Antonov et al., 2008). For host species that have evolved advanced egg discrimination strategies, the recognition of alien eggs should be relatively simple and error-free, such that rapid decision-making and rejection are expected. However, for hosts with weak recognition abilities, hosts need to repeatedly examine eggs and only begin attempting rejection when motivation exceeds a certain threshold, as a way to minimise the risk of recognition errors. We suggest that the low rate of egg rejection in the short period, for the Crested Myna, is largely due to the repeated perception of alien eggs during the decision-making process.
Hosts mostly use eggshell characteristics (colour, patterning, etc.) as recognition cues of alien eggs, but a few species use also egg size to clue alien eggs (Ye et al., 2022). However, it is not clear whether the rejecting eggs that differ in size has evolved as a consequence of parasitism selection pressures. For example, these studies have either found a mixed effect of egg size and egg colour/patterning on egg recognition (Mason and Rothstein, 1986; Tosi-Germán et al., 2020) or suggested that egg size affects the ejection rate but not host egg recognition per se (Roncalli et al., 2016). Roncalli et al. (2016) found that Western Bonelli’s Warbler (Phylloscopus bonelli) exhibits a significantly lower ejection rate with large model eggs than with small model eggs. However, the authors argued that the size of eggs was not a clue to the recognition of eggs of this small species, namely Bonelli’s Warbler, as this bird more often chose to abandon nests containing large model eggs and ejected large eggs for a significantly longer period compared to that with small eggs. This is possibly because the ejection of large model eggs, which has a large rejection cost across multiple attempts, is limited by the size of the parent bird. Similarly, our results showed significantly lower ejection rates with large eggs than with small and medium eggs, but we suggest that the Crested Myna uses egg size as an egg recognition cue to reject foreign eggs, mainly because of the following reasoning: (1) the Crested Myna is fully capable of rejecting Japanese Quail eggs, but the Crested Myna accepted all Japanese Quail eggs and did not abandon the nest; (2) none of the large eggs were rejected by Crested Mynas, which might be in accordance with the parental investment theory, which suggests that, because of their higher fitness prospect (Perrins, 1965), parents prefer larger eggs (Tinbergen, 1951). In addition, Crested Mynas rejected 58.3% of the small experimental eggs, which supports the idea that this species rejects eggs using egg size as a recognition cue. However, the egg size may not be the only cue for egg rejection; next, we will continue to explore the influence of other factors on egg recognition, such as nest brightness.
Two main hypotheses have been used to explain the cognitive mechanisms underlying egg recognition in host birds as follows: (1) true or template recognition, where the host uses its own eggs as a template (innate ability and/or through learning) to accurately identify and reject alien eggs; and (2) recognition by discordancy, where the host rejects the eggs that differ in appearance from the more abundant egg type in the nests. Thus, species following this rule do not need an innate template or learning the appearance of their own eggs for detecting parasitic eggs (Rothstein, 1974; 1975; Bán et al., 2013). Currently, a few birds use discordancy recognition, and this mechanism usually coexists with true or template recognition (Yang et al., 2014a; Yang et al., 2014b; Wang et al., 2015). Our study demonstrates that the cognitive mechanism of egg recognition in the Crested Myna is discordancy recognition and that foreign eggs are recognised through a comparison of differences in egg size in the nest, as the Crested Myna rejects a significantly higher proportion of small eggs in the nest than large eggs, even when their own eggs are the scarcer egg type.
In addition, our results showed that the rejection rate of small-sized Magpie Robin nestlings was 100%, and this was 33.3% for large-sized Magpie Robin nestlings and 0% for large-sized Grey-backed Starling nestlings, within the nests of the Crested Myna. Therefore, it is quite likely that the Crested Myna also uses nestling size as a cue for nestling recognition. There is a big difference in body colour and begging sounds between Oriental Magpie Robin’s nestlings and Crested Myna’s nestlings (Fig. 3), but mynas accepted Oriental Magpie Robin’s nestlings, which suggests that Crested Mynas may not discriminate their own nestlings using visual and/or auditory cues. However, the death and disappearance of heterospecific nestling can also be incurred by difference of predation rate and/or food type preferences between the nestling and the fosters. In addition, there are other factors to be considered like survival rates depending on chick ages and environmental conditions (e.g., the length of the beak split, nest temperatures and humidity). Therefore, the nestling size may not be the only cue for nestling rejection. Unfortunately, the sample size of our experiment was relatively small considering ecological ethics and other issues, and other experimental tools will be considered in the future to continue exploring the link between nestling size factors and parent–nestling discrimination.
5.
Conclusion
In summary, our experimental results support the hierarchical correlation between nest sanitation and egg rejection behaviour proposed by Moskát et al. (2003). Our study provides clear evidence that the Crested Myna uses egg size as a cue for recognition rather than egg colour or egg patterning. Our study suggests that the cognitive mechanism of the Crested Myna is based on the discordancy recognition of egg size as a recognition cue. It is highly likely that parent–nestling recognition is also based on size as a recognition cue, but the sample size of our experiment was small given ecological, ethical, and other issues. In addition, our study found a temporal delay in egg rejection behaviour in the Crested Myna; therefore, we suggest that the periodicity of egg rejection experiments could be appropriately extended, especially for species with relatively low egg recognition ability.
Authors’ contribution
WL conceived and designed this study. JL, FZ and YL carried out field experiments. JL performed data analyses and drafted the manuscript, and WL revised and improved the manuscript. All authors read and approved the final manuscript.
Ethical statements
The experiments comply with the current laws of China. Fieldwork was performed with permission from villages of Ding’an and Wenchang, Hainan, China. Experimental procedures were in accordance with the Animal Research Ethics Committee of Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (no. HNECEE-2012-001).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We are grateful to local people in villages of Ding’an and Wenchang, Hainan, for their help and cooperation.
Abolins-Abols, M., Hanley, D., Moskát, C., Grim, T., Hauber, M.E., 2019. Anti-parasitic egg rejection by great reed warblers (Acrocephalus arundinaceus) tracks differences along an eggshell color gradient. Behav. Process. 166, 103902.
Antonov, A., Stokke, B.G., Moksnes, A., Røeskaft, E., 2008. Getting rid of the cuckoo Cuculus canorus egg: why do hosts delay rejection? Behav. Ecol. 19, 100-107.
Bán, M., Moskát, C., Barta, Z., Hauber, M.E., 2013. Simultaneous viewing of own and parasitic eggs is not required for egg rejection by a cuckoo host. Behav. Ecol. 24, 1014-1021.
Craig, A., Feare, C., 1998. Starlings and Mynas. Christopher Helm, London.
Davies, N.B., 2000. Cuckoos, Cowbirds and Other Cheats. T & AD Poyser, London.
Davies, N.B., Brooke, M.L., 1989. An experimental study of co-evolution between the cuckoo Cuculus canorus and its hosts. I. Host egg discrimination. J. Anim. Ecol. 58, 207-224.
Davies, N.B., Brooke, M.De.L, Kacelnik, A., 1996. Recognition errors and probability of parasitism determine whether reed warblers should accept or reject mimetic cuckoo eggs. Proc. R. Soc. B 263, 925-931.
Feng, C., Yang, C., Liang, W., 2019. Nest sanitation facilitates egg recognition in the common tailorbird, a plaintive cuckoo host. Zool. Res. 40, 466-470.
Forsman, J.T., Martin, T.E., 2009. Habitat selection for parasite-free space by hosts of parasitic cowbirds. Oikos 118, 464-470.
Goguen, C.B., Mathews, N.E., 1996. Nest desertion by blue-gray gnatcatchers in association with brown-headed cowbird parasitism. Anim. Behav. 52, 613-619.
Grim, T., 2006. The evolution of nestling discrimination by hosts of parasitic birds: why is rejection so rare? Evol. Ecol. Res. 8, 785-802.
Grim, T., Honza, M., 2001. Differences in behaviour of closely related thrushes (Turdus philomelos and T. merula) to experimental parasitism by the common cuckoo Cuculus canorus. Biologia 56, 549-556.
Guigueno, M.F., Sealy, S.G., 2009. Nest sanitation plays a role in egg burial by yellow warblers. Ethology 115, 247-256.
Guigueno, M.F., Sealy, S.G., 2010. Clutch abandonment by parasitized yellow warblers: Egg burial or nest desertion? Condor 112, 399-406.
Guigueno, M.F., Sealy, S.G., 2012. Nest sanitation in passerine birds: implications for egg rejection in hosts of brood parasites. J. Ornithol. 153, 35-52.
Hanley, D., López, A.V., Fiorini, V.D., Reboreda, J.C., Grim, T., Hauber, M.E., 2019. Variation in multicomponent recognition cues alters egg rejection decisions: A test of the optimal acceptance threshold hypothesis. Philos. T. Roy. Soc. B 374, 20180195.
Holveck, M.J., Doutrelant, C., Guerreiro, R., Perret, P., Gomez, D., Grégoire, A., 2010. Can eggs in a cavity be a female secondary sexual signal? Male nest visits and modelling of egg visual discrimination in blue tits. Biol. Lett. 6, 453-457.
Honza, M., Cherry, M.I., 2017. Egg characteristics affecting egg rejection. In: Soler, M. (Ed.), Avian Brood Parasitism. Switzerland Springer International Publishing, Cham.
Honza, M., Kuiper, S.M., Cherry, M.I., 2005. Behaviour of African turdid hosts towards experimental parasitism with artificial red-chested cuckoo Cuculus solitarius eggs. J. Avian Biol. 36, 517-522.
Hosoi, S., Rothstein, S.I., 2000. Nest desertion and cowbird parasitism: evidence for evolved responses and evolutionary lag. Anim. Behav. 59, 823-840.
Kilner, R.M., 2006. The evolution of egg colour and patterning in birds. Biol. Rev. 81, 383-406.
Krüger, O., 2007. Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Philos. T. Roy. Soc. B 362, 1873-1886.
Langmore, N.E., Hunt, S., Kilner, R.M., 2003. Escalation of a coevolutionary arms race through host rejection of brood parasitic young. Nature 422, 157-160.
Li, D., Li, X., Zhang, Y., Guan, S., Ruan, Y., 2020. Contrasting effects of egg size and appearance on egg recognition and rejection response by Oriental reed warblers. Ecol. Evol. 10, 10508-10519.
Li, Q., Bi, J., Wu, J., Yang, C., 2021. Impact of nest sanitation behavior on hosts’ egg rejection: an empirical study and meta-analyses. Curr. Zool. 67, 683-690.
Liu, J., Liang, W., 2021. The breeding ecology of the crested myna Acridotheres cristatellus on tropical Hainan Island. Ornithol. Sci. 20, 83-92.
Liu, C., Ye, P., Cai, Y., Quan, R., Yang, C., 2021. Persistent fine-tuning of egg rejection based on parasitic timing in a cuckoo host even after relaxation of parasitism pressure. Behav. Process. 193, 104532.
Lotem, A., Nakamura, H., Zahavi, A., 1995. Constraints on egg discrimination and cuckoo host coevolution. Anim. Behav. 49, 1185-1209.
Luro, A.B., Hauber, M.E., 2017. A test of the nest sanitation hypothesis for the evolution of foreign egg rejection in an avian brood parasite rejecter host species. Sci. Nat. 104, 14.
Luro, A.B., Igic, B., Croston, R., López, A.V., Shawkey, M.D., Hauber, M.E., 2018. Which egg features predict egg rejection responses in American robins? Replicating Rothstein's 1982 study. Ecol. Evol. 8, 1673-1679.
Mason, P., Rothstein, S.I., 1986. Coevolution and avian brood parasitism: cowbird eggs show evolutionary response to host discrimination. Evolution 40, 1207-1214.
Møller, A.P., Diaz, M., Liang, W., 2016. Brood parasitism and proximity to human habitation. Behav. Ecol. 27, 1314-1319.
Moskát, C., Honza, M., 2002. European cuckoo Cuculus canorus parasitism and host’s rejection behaviour in a heavily parasitized great reed warbler Acrocephalus arundinaceus population. Ibis 144, 614-622.
Moskát, C., Székely, T., Kisbenedek, T., Karcza, Z., Bártol, I., 2003. The importance of nest cleaning in egg rejection behaviour of great reed warblers Acrocephalus arundinaceus. J. Avian Biol. 34, 16-19.
Noh, H.J., Gloag, R., Langmore, N.E., 2018. True recognition of nestlings by hosts selects for mimetic cuckoo chicks. Proc. R. Soc. B 285, 20180726.
Peer, B.D., 2017. Nest sanitation does not elicit egg ejection in a brown-headed cowbird host. Anim. Cognition 20, 371-374.
Peer, B.D., Sealy, S.G., 2004. Correlates of egg rejection in hosts of the brown-headed cowbird. Condor 106, 580-599.
Perrins, C.M., 1965. Population fluctuations and clutch-size in the great tit Parus major L. J. Anim. Ecol. 34, 601-647.
Polačikova, L., Honza, M., Procházka, P., Topercer, J., Stokkes, B., 2007. Color characteristics of the blunt egg pole: cues for recognition of parasitic eggs as revealed by reflectance spectrophotometry. Anim. Behav. 74, 419-427.
Požgayová, M., Procházka, P., Polačiková, L., Honza, M., 2011. Closer clutch inspection-quicker egg ejection: timing of host responses toward parasitic eggs. Behav. Ecol. 22, 46-51.
Rodríguez-Gironés, M.A., Lotem, A., 1999. How to detect a cuckoo egg: a signal-detection theory model for recognition and learning. Am. Nat. 153, 633-648.
Roncalli, G., Ibáñez-Álamo, J.D., Soler, J.J., 2016. Size and material of model parasitic eggs affect the rejection response of Western Bonelli’s warbler Phylloscopus bonelli. Ibis 159, 113-123.
Rothstein, S.I., 1974. Mechanisms of avian egg recognition: possible learned and innate factors. Auk 91, 796-807.
Rothstein, S.I., 1975. Mechanisms of avian egg-recognition: do birds know their own eggs? Anim. Behav. 23, 268-278.
Rothstein, S.I., 1990. A model system for coevolution: avian brood parasitism. Ann. Rev. Ecol. Syst. 21, 481-508.
Sadam, A., 2022. Breeding ecology and anti-brood parasitism strategy of the Common Myna in Pakistan. Doctoral Thesis. Hainan Normal University, Haikou.
Sato, N.J., Tokue, K., Noske, R.A., Mikami, O.K., Ueda, K., 2010. Evicting cuckoo nestlings from the nest: a new anti-parasitism behaviour. Biol. Lett. 6, 67-69.
Sealy, S.G., Neudorf, D.L., 1995. Male northern orioles eject cowbird eggs: implications for the evolution of rejection behavior. Condor 97, 369-375.
Segura, L.N., Sallo, F.D., Mahler, B., Reboreda, J., 2016. Red-crested Cardinals use color and width as cues to reject Shiny Cowbird eggs. Auk 133, 308-315.
Soler, M., 2014. Long-term coevolution between avian brood parasites and their hosts. Biol. Rev. 89, 688-704.
Soler, M., Fernández-Morante, J., Espinosa, F., Martín-Vivaldi, M., 2012. Pecking but accepting the parasitic eggs may not reflect ejection failure: the role of motivation. Ethology 118, 662-672.
Soler, M., Ruiz-Raya, F., Roncalli, G., Ibáñez-Álamo, J.D., 2017. Relationships between egg-recognition and egg-ejection in a grasp-ejector species. PLoS One 12, e0166283.
Spottiswoode, C.N., Stevens, M., 2010. Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs. P. Natl. Acad. Sci. USA 107, 8672-8676.
Su, T., Yang, C., Chen, S., Liang, W., 2018. Does nest sanitation elicit egg rejection in an open-cup nesting cuckoo host rejecter? Avian Res. 9, 27.
Tinbergen, N., 1951. The Study of Instinct. Clarendon Press, Oxford.
Tokue, K., Ueda, K., 2010. Mangrove Gerygones Gerygone laevigaster eject Little Bronze-cuckoo Chalcites minutillus hatchlings from parasitized nests. Ibis 152, 835-839.
Tosi-Germán, R.A., Tassino, B., Reboreda, J.C., 2020. Female and male rufous horneros eject shiny cowbird eggs using a mental template of the size of their own eggs. Behav. Process. 178, 104152.
Wang, L., Yang, C., Møller, A.P., Liang, W., Lu, X., 2015. Multiple mechanisms of egg recognition in a cuckoo host. Behav. Ecol. Sociobiol. 69, 1761-1767.
Welbergen, J.A., Davies, N.B., 2009. Strategic variation in mobbing as a front line of defense against brood parasitism. Curr. Biol. 19, 235-240.
Yang, C., Møller, A.P., Røskaft, E., Moksnes, A., Liang, W., Stokke, B.G., 2014. Reject the odd egg: Egg recognition mechanisms in parrotbills. Behav. Ecol. 25, 1320-1324.
Yang, C., Wang, L., Cheng, S.J., Hsu, Y.C., Liang, W., Moeller, A.P., 2014. Nest defenses and egg recognition of yellow-bellied prinia against cuckoo parasitism. Naturwissenschaften 101, 727-734.
Yang, C., Chen, M., Wang, L., Liang, W., Møller, A.P., 2015a. Nest sanitation elicits egg discrimination in cuckoo hosts. Anim. Cognition 18, 1373-1377.
Yang, C., Wang, L., Liang, W., Møller, A.P., 2015b. Nest sanitation behavior in hirundines as a pre-adaptation to egg rejection to counter brood parasitism. Anim. Cognition 18, 355-360.
Yang, C., Wang, L., Chen, M., Liang, W., Møller, A.P., 2015c. Nestling recognition in red-rumped and barn swallows. Behav. Ecol. Sociobiol. 69, 1821-1826.
Ye, P., Cai, Y., Wu, N., Yao, X., Li, G., Liang, W., et al., 2022. Egg rejection based on egg size recognition as a specific strategy against parasitic cuckoos. Curr. Zool. 69, 156-164.
Yi, T., 2020. Coevolution between large hawk cuckoos and its hosts. Doctoral Thesis. Hainan Normal University, Haikou.
Zhang, J., Møller, A.P., Yan, D., Li, J., Deng, W., 2021. Egg rejection changes with seasonal variation in risk of cuckoo parasitism in Daurian redstarts, Phoenicurus auroreus. Anim. Behav. 175, 193-200.
Zheng, G., 2017. A Checklist on the Classification and Distribution of the Birds of China. 3rd ed. Science Press, Beijing.
Table
3.
Results of experiments on egg recognition mechanisms and parent-nestling recognition in the Crested Myna (L refers to large and S to small size groups).
Experiment operation
All accepted
S rejected
L rejected
S and L rejected
Three quail eggs (L) + one myna (S)
7
4
4
1
Three quail eggs (L) + one cockatiel (S)
4
6
1
4
Three cockatiel eggs (S) + one myna (L)
13
6
0
2
Two quail eggs (L) + two cockatiel (S)
5
7
1
1
Myna nestlings (S) + one starling (L)
3
0
0
0
Myna nestlings (S) + one magpie robin (L)
2
0
1
0
Myna nestlings (L) + one magpie robin (S)
0
4
0
0
Myna nestlings (S) + one myna (L)
4
0
0
0
Myna nestlings (L) + one myna (S)
4
0
0
0
The number in the table refers to the number of nests.


 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds