School of Life Sciences, Nanchang University, Nanchang, 330031, China
b.
Faculty of Agricultural and Environmental Sciences, McGill University, QC, Canada
c.
Jinhui Liquor Company Limited, Longnan, 742300, China
d.
Guangdong Maoming Health Vocational College, Maoming, 525000, China
e.
Jiangxi Province Key Laboratory of Watershed Ecosystem Change and Biodiversity, Center for Watershed Ecology, Institute of Life Science and School of Life Sciences, Nanchang University, Nanchang, 330031, China
Urbanization is currently considered one of the most rapid types of global environmental change. Urban habitats are biotically and abiotically different from their rural areas, i.e., the ambient temperature, predator, and food availability. These novel challenges create new selection pressures, which allow one to investigate eco-evolutionary responses to contemporary environmental change. A total of 118 breeding nests were monitored for nest predation in both urban and rural areas from 2018 to 2020. We used environmental factors from urban and rural areas and behavioral data from 439 Chinese Blackbird (Turdus mandarinus) valid incubation days to understand the impact of urbanization on the incubation behavior of blackbirds and its adaptation mechanism to the urban environment. Cities have warmer ambient temperatures and lower predation pressures than rural areas. Urban blackbirds chose the incubation strategy with shorter and more bouts, while rural blackbirds selected the incubation strategy with longer and fewer bouts. The plasticity of incubation behavior of urban blackbirds was higher than that of rural areas, and the range of egg temperature was also higher than that of rural areas. In addition, incubation temperature and the number of bouts per day were the key factors affecting the day survival rate of blackbirds, and the hatching rate of urban blackbirds was higher than that of rural blackbirds. Our results provide evidence for behavioral shifts in blackbirds during adaptation to urbanization and support the central role of behavioral adaptation in the successful colonization of new environments by wildlife. These help us understand the behavioral characteristics required for wildlife to live in cities and the urban adaptors faced environmental pressures.
Urbanization is one of the most extreme forms of environmental alteration, posing a major threat to biodiversity and altering the ecosystem on which human civilization depends (Kurucz et al., 2021; Cronin et al., 2022; Curtis et al., 2022). The environmental structuring of cities so clearly differs from natural habitats, with reduced natural vegetation, elevated levels of human disturbance, distinct predator compositions, and increased light, noise, and heavy metal pollution, as well as warmer temperatures (Vincze et al., 2021; Batisteli et al., 2022; Bailey et al., 2022). On the other side, urbanization results in an alternation of high-density human settlements, industrial zones, and buildings, frequently intermixed with managed green spaces or natural habitat remnants (McIntyre et al., 2001; Grimm et al., 2008; González-Lagos et al., 2021). This environment with strong human disturbance has accelerated phenotypic diversification and impaired reproductive success for wild animals (Corsini et al., 2021; Smith et al., 2021). As an essential component of the urban ecosystem, birds are highly sensitive and susceptible to respond significantly to environmental change (Bailey et al., 2022). Therefore, birds are an excellent model system for assessing urban-driven evolutionary shifts (Havlíček et al., 2021).
At the moment, an increasing number of ecological research has concentrated on how birds successfully settle in urban environments (Corsini et al., 2021). Birds have to adjust their reproductive strategies to urban environments due to brand-new selection pressures carried on by urbanization (Zhou et al., 2020; Bujan et al., 2021; Vincze et al., 2021). Assimilation lighting in cities may stimulate gonad development in birds, increasing their probability of expressing reproductive behavior (Dawson et al., 2001), leading to birds mating and nesting ahead of time. Furthermore, the stable and long-term availability of artificial food in cities stimulated birds to nest early (Crates et al., 2016). When it comes to incubation and nestling periods, however, urban birds are likely to be more affected (Heppner and Ouyang, 2021; Hope et al., 2021; Vincze et al., 2021), such as urban birds exhibited smaller clutches and brood sizes compared to rural avian populations (Bailly et al., 2016; Kettel et al., 2018; Seress et al., 2020).
Incubation is an essential component of the breeding cycle in birds, and parental effort during incubation can have substantial solid consequences on the reproductive success of the pair (Deeming and Gray, 2016). The incubation quality also influences reproductive success in birds that care for their eggs. Avian embryo requires a narrow species-specific thermal range for optimal development in which incubation temperature outside the limits results in embryonic development abnormalities (Carvalho et al., 2020; Costa et al., 2020; Grimaudo et al., 2020). Even slight alterations in the ambient temperature can impact the phenotypic growth rates, immunological function, and survival of offspring (Ardia et al., 2010; Merrill et al., 2020).
As a result, incubating birds must trade off leaving the nest to forage with staying on the nest to incubate, i.e., incubating birds should maintain optimal embryo development temperatures for developing embryos while meeting their energetic demands (Webb and Cooperpreston, 1989; Heppner and Ouyang, 2021). Incubation behavior manifests this trade-off and is heavily influenced by the microenvironment (Kleindorfer et al., 2021), food availability (Nord and Cooper, 2020; Ryeland et al., 2021), attentiveness of the nest (Heppner and Ouyang, 2021) and nest predation risk (Yoon et al., 2017). More and shorter bouts of recess can lead to lower nest attentiveness when adults are exposed to urban habitats with warmer ambient temperatures (Batisteli et al., 2021; Ton et al., 2021). In rural areas with higher nest predation (Conway and Martin, 2000), there may be a trade-off between increasing nest activity to maintain endogenous reserves for the incubating parents and minimizing nest activity to minimize nest predation risk (Fontaine and Martin, 2006). For example, birds will leave the nest fewer times and for more extended periods in rural habitats (Conway and Martin, 2000; Fontaine and Martin, 2006). Additionally, the food resources available for carnivorous birds in urban, including insects and worms, are significantly lower (Fenoglio et al., 2021). Low-quality food would increase the energetic costs of foraging trips, leading to more frequent and prolonged recesses (Londono et al., 2008).
Incubation is also an extremely time-expending and energy-consuming behavior (Thomson et al., 1998), so female-only incubation is exceptionally constrained (Deeming, 2002, Nord and Williams, 2015) because of their high energetic investments in gametes and parental care (Nord and Nilsson, 2011; Plaza et al., 2018). The effect of urbanization on bird reproduction has been extensively investigated. However, urbanization has not been systematically studied in terms of how it affects the hatching strategies of birds. Additionally, further research should be conducted on the ways in which urbanization may affect bird reproduction and succession by modulating incubation strategy. Accordingly, the aim of this study is to evaluate the effects of urbanization on bird hatching countermeasures by studying the Chinese Blackbird (Turdus mandarinus).
The Chinese Blackbird is a female-only incubation bird that can adapt to a multitude of environments and is one of the most common urban and natural habitats breeding birds (Wang et al., 2009). In recent decades, the Chinese Blackbird has begun to colonize and successfully reproduce in and around major young cities (Han et al., 2019; Luscier, 2022). The focus of this research was to describe incubation behavioral changes among Chinese Blackbirds in urban and rural environments. Chinese Blackbirds made the following predictions about the significance of incubation behavioral adaptation in the urban and rural environments: 1) in the presence of lower environmental stresses, urban individuals expend less energy to reach higher production, resulting in improved reproductive success and fitness; and 2) in urban habitats, the enhanced plasticity of incubation behavior enables citizens to better adapt to urbanized landscapes.
2.
Methods
2.1
Study areas
In the breeding season (March to July) from 2018 to 2020, this study focused on two locations, including Nanchang (urban) and Huangjinpu in Yugan County (rural), China. The primary types of land at the urban site (115°47′12″–115°38′34″ E, 28°39′4″–28°40′19″ N) were artificial habitats (Han et al., 2019), while the rural site (116°13′48″–116°54′24″ E, 28°21′36″–29°3′24″ N) comprised farmland and natural woodland. Both sites were subtropical humid monsoon climates with Chinese Blackbirds breeding every year. Blackbirds have a breeding density of 31–32 breeding pairs per square kilometer in urban areas, while 1–2 breeding pairs per square kilometer in rural areas.
2.2
General field methods
Each study region was visited at seven days intervals to locate breeding pairs and nests at the beginning of each breeding season. The observations were carried out with 10 × 42 binoculars (Swarovski) from the urban and rural sites. The main ways to find the nest site include that: 1) the bird was observed carrying nest material to the vegetation belt, 2) the bird was spotted leaving a potential nest tree, or 3) nest was directly observed in trees. When located, a nest was numbered and its position recorded with the Google Earth Pro program (Version 7.1.8, Google, 2020). We increased the frequency of visits to three days during the laying period. Visiting during the laying period for most nests permitted us to detect and judge whether incubation was starting (presence of uncovered and warm eggs or direct observation of an incubating female). Each nest was also visited every three days in incubation periods to determine hatch date, nest fate, clutch size, and brood size. We defined the hatching termination as until the last egg hatched or a new hatchling was absent for two consecutive visits. We estimated days-to-hatch and initiation date based on the number of eggs laid if nests were found during egg-laying (assuming one egg laid per day) and by back-calculating from the known hatch date using standard incubation duration). Finally, we retrieved the data logger (see below) when the breeding was completed. We excluded nests that failed to breed due to predation, and then calculated hatching rates using the brood size/clutch size per nest. Because differentiating between fertilized and unfertilized eggs during breeding can easily lead to nest abandonment by birds, and because egg infertility is usually random in natural populations (Zhang et al., 2003), this case was not excluded in our analysis.
The passive infrared camera trap (LTL Acorn 6210 MC infrared digital cameras) was used to assess egg predation to monitor the breeding nest. To reduce overexposure and unwanted triggering events induced by the sunrise and sunset, each camera was adjusted to face north or south to the extent possible (Zhu et al., 2021). In order to capture three images plus one 10-s video per trigger, the camera was set to medium sensitivity. Each camera was equipped with 12 batteries and a 64 GB memory card. At intervals of three days, we replaced the memory card and battery in the camera. If necessary, the camera angle was adjusted to face the nests' most explicit view area during every routine check. A thorough analysis of the photo and video data has been conducted to determine the species and the number of egg predators.
2.3
Temperature data collections
We recorded nest temperature at Chinese Blackbird nests using the Elitech temperature data logger (RC-4HC, accuracy ± 0.5 ℃, Elitech Technology, Inc.). The data logger was programmed to record temperature (accuracy ± 0.5 ℃) every 2 min. When the egg was detected, a data logger was carefully placed in the nest and ensured its probe was anchored to the clutch's center, surrounded by the eggs (Croston et al., 2020). To measure ambient temperature in the nest, another data logger was placed on the tree trunk supported by the nests, fixed with scotch tape, recording local ambient temperature every 2 min.
2.4
Incubation behavior
We used R package incR to analyze incubation behavior (Capilla-Lasheras, 2018). Using the method implemented in incR, we exploited both variations in incubation and ambient temperatures to calculate the presence (on bouts: times when a female is feeding or incubating the clutch) or absence (off bouts: times when a female is foraging or conducting other self-maintenance activities outside of the nest) of an incubating individual. Then incRenv function was used to automatically assign an environmental temperature to every incubation temperature observation, information required by the incRscan function to score incubation. The IncRact function was applied to calculate onset (first off-bout in the morning) and offset (last on-bout in the evening). We also calculated day nest attentiveness (as the percentage of day time spent for each nest), day incubation temperature, and nightly incubation temperature, using the incRatt function in the incR package. Finally, we employed the incRbout function to calculate the number of on-bouts and off-bouts per day and their average duration, starting time, duration, starting temperature and final temperature for every on-bout and off-bout detected by incRscan.
2.5
Statistical analyses
In this study, the normality of the distributions was assessed using the Kolmogorov-Smirnov test, with the significance level set at 0.05 (Lilliefors, 1987). In both urban and rural areas, the coefficient of variation was used to estimate the level of dispersion, average off-bout duration and average incubation duration. A simple linear regression analysis was conducted to evaluate the Julian date association with ambient mean temperature, average incubation duration, and nest attentiveness, and the ambient mean temperature relationship with egg temperature and nest attentiveness. The Simba package was then applied to test whether the regression coefficients or intercepts differed significantly at the 0.05 level. Uncertainties associated with each regression were characterized by the 95% confidence interval estimated from the standard error of each regression.
Pearson correlation test was performed to assess the correlation among the incubation parameters (Appendix Table S1). To reduce over-fitting caused by redundant variables, we selected the most explanatory and uncorrelated coefficients (|r| > 0.70; Han et al., 2019). After selecting the variable that provided the most meaningful biological interpretation, the remaining variables were removed from further analysis, resulting in a set of seven variables (including day egg temperature, day egg temperature variation, night egg temperature variation, number of bouts, incubation total, offset and onset).
We used a binomial generalized linear regression model to predict various locations (rural/urban) and to evaluate the influence of seven incubation factors. The effect of seven incubation factors on the nest day survival rate (DSR) of Chinese Blackbirds was also estimated using a negative binomial regression model. DSR, the probability that a nest will survive a single day, was calculated using the known-fate model with the RMark package. All final models met assumptions of normality and homoscedasticity of residual errors. Significance was taken at a 0.05 level. Data exploration again showed a non-independent temporal structure of residuals, and models were built accordingly. We selected the most informative model based on Akaike's information criterion (AIC). Better fitting models were having lower AIC values via the MuMIn package. A candidate set of models was first produced with all possible parameter subsets, which were then fitted to the data and ranked by ΔAICc values (the difference between each model's AICc and AICc minimum, that of the "best" model). Top models with ΔAICc < 2 were reported in the tables.
We analyzed the final selected models and reported their R2 values via the MuMIn package. All analyses were conducted in R software 4.0.6 (R Core Team, 2021). All statistical tests used two-tailed p values with a threshold of 0.05.
3.
Results
3.1
Environmental factors differing between urban and rural regions
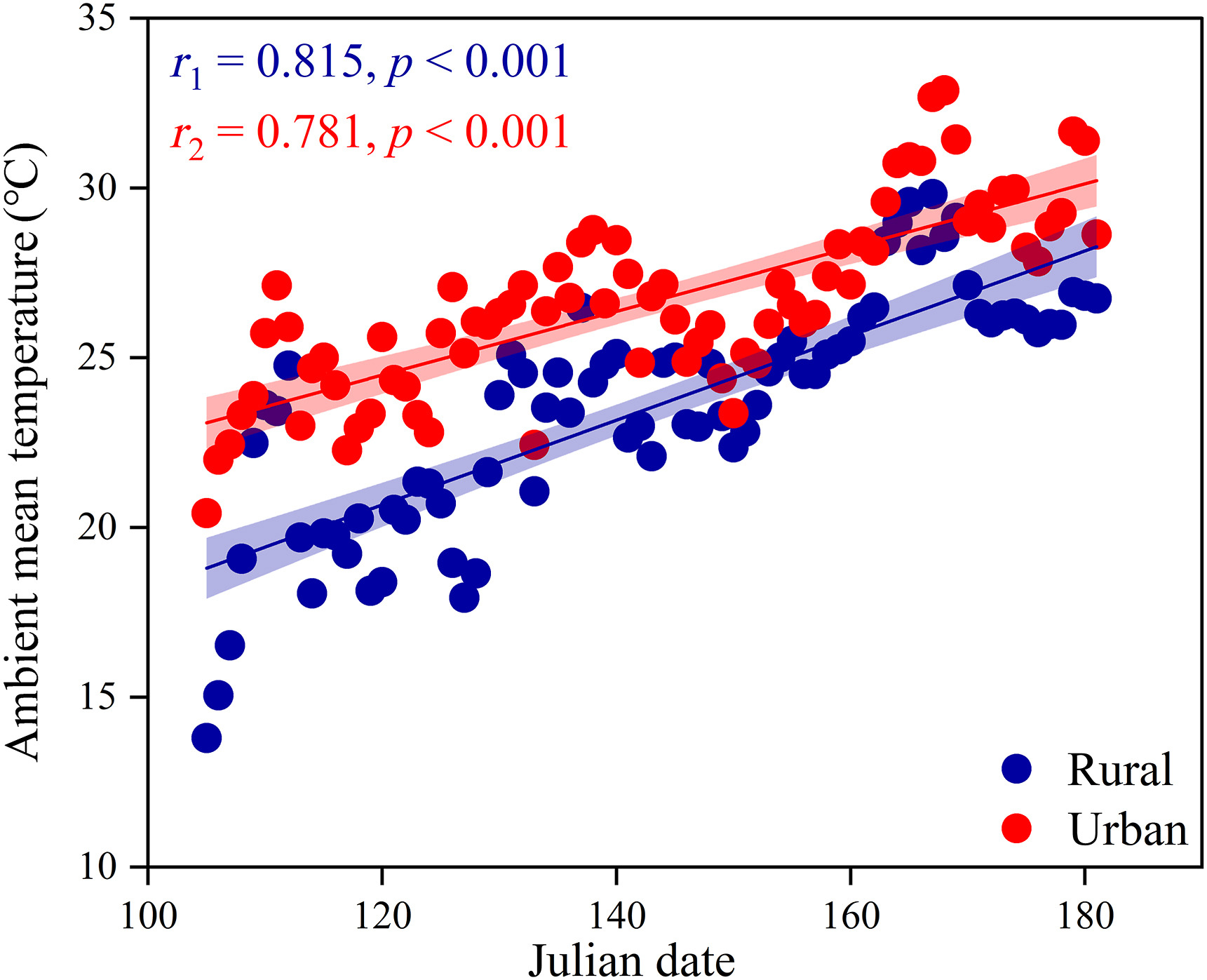
For the impact of COVID-19, predation and nest abandonment, the final data collected was 439 valid incubation days, including 108 days for the rural population and 331 days for the urban population. From April to July, the ambient mean temperature in the city is 26.65 ± 2.69 ℃, whereas it is 23.53 ± 3.42 ℃ in the rural areas. Our results showed that the ambient mean temperature in urban areas is significantly higher than in rural areas (t = 6.28, df = 144, p < 0.001). The ambient mean temperature of the two regions increased continuously and significantly (urban: r = 0.78, p < 0.001; rural: r = 0.82, p < 0.001; Fig. 1). There was no significant difference between the estimated and slope regression line derived urban and rural temperature (t = 0.30, df = 73, p = 0.763).
Figure
1.
The difference of urban and rural ambient temperature in breeding season. Urban site and females are in red points, and rural site and females are in blue points. Linear regressions with 95% confidence intervals are shown within each panel, and lines represent the best fit from a linear model. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
From 2018 to 2020, we monitored 118 breeding nests. There were 90 breeding nests in urban areas; 14 of them had been invaded by predators, accounting for 15.56% of the total. While in the 28 nests in rural areas, 12 had been invaded by predators, accounting for 42.86% of the total. Chi-square test shows that the nest predation rate in urban areas was significantly lower than that in rural areas (χ2 = 12.76, df = 1, p < 0.001). Then, we analyzed the types of nest predators detected by infrared cameras, and we found that urban predators were divided into the sky and the ground predators. The former includes Eurasian Jay (Garrulus glandarius), Red-billed Blue Magpie (Urocissa erythroryncha), Great Tit (Parus major), and Long-tailed Shrike (Lanius schach) in the sky, while the latter consists of Crimson Snake (Dinodon rufozonatum), Rattus Tanezumab (Rattus flavipectus) and Stink Rat Snake (Elaphe carinata). Rural predators, however, come mainly from the ground including Crimson Snake and Stink Rat Snake.
3.2
Incubation parameters varing between urban and rural birds
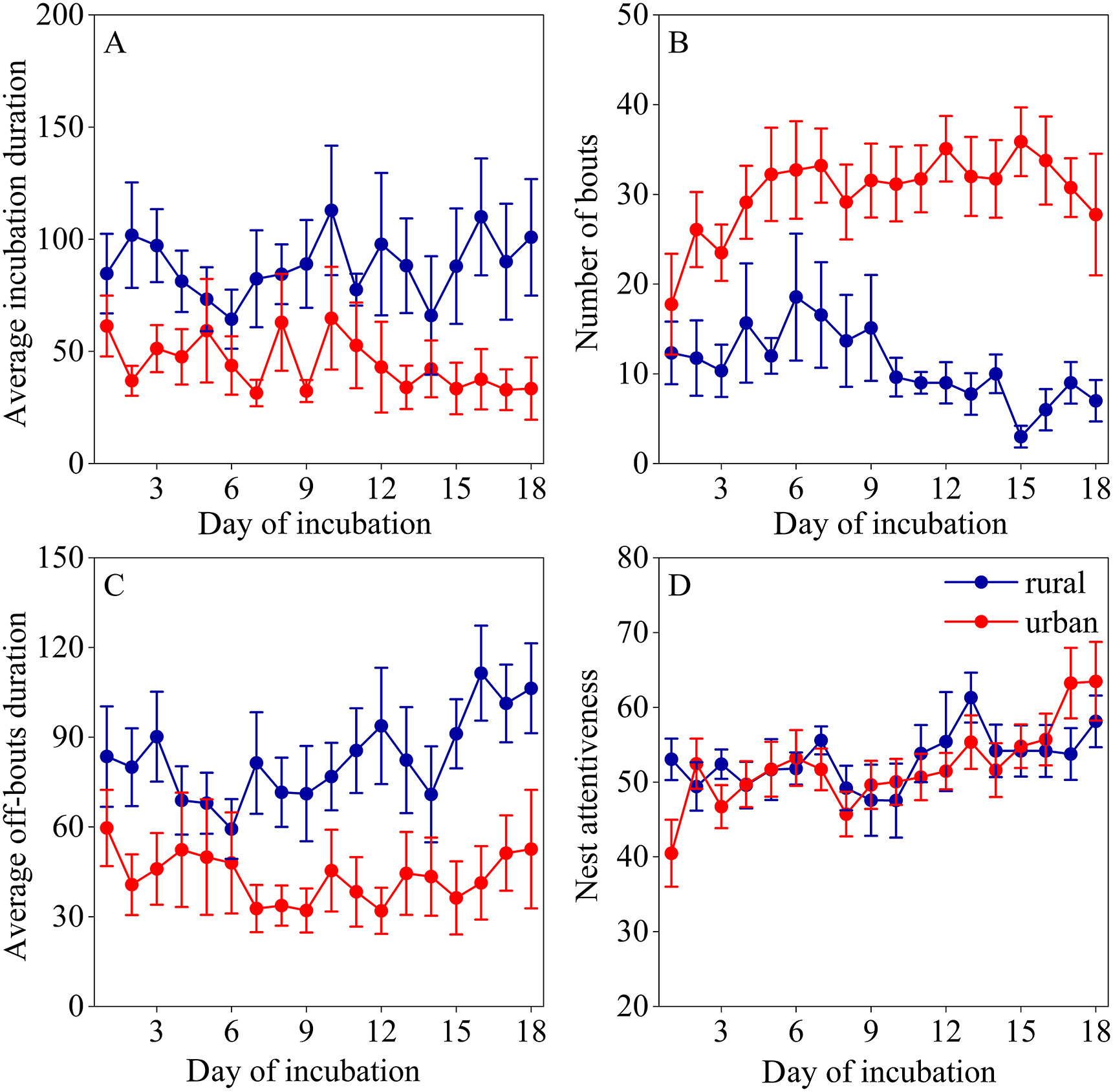
Chinese Blackbirds in urban areas had an average incubation duration of 38.41 ± 43.56 min, with a day total incubation duration of 710.54 ± 133.78 min. The Chinese Blackbird's total day incubation duration was 710.97 ± 119.41 min in rural regions, with an average incubation duration of 75.70 ± 29.40 min (Table 1). Urban females had an average shorter incubation duration than rural females (Fig. 2A) with significant difference (Table 1). Urban females had higher number of bouts per day than rural females (Fig. 2B) with significant difference (z = −4.30, df = 38, p < 0.001). As the incubation period progressed, rural females decreased the number of bouts while urban females' number of bouts was maintained (Fig. 2B). The average off-bouts duration increased among rural females as the incubation period progressed (Fig. 2C). Additionally, the nest attentiveness (urban: 49.41 ± 8.31; rural: 52.03 ± 8.78) and total incubation duration of urban females were not significantly different from rural females (Table 1). Rural and urban females nest attentiveness was also maintained as the incubation period progressed (Fig. 2D).
Table
1.
Descriptive statistics and analysis of variance for incubation parameters in urban and rural areas.
Incubation parameters
Rural (n = 10)
Urban (n = 29)
T/Mann-Whitney test
Mean
SD
Mean
SD
t/Z
p
Day egg temperature
23.79
1.08
22.97
2.95
1.27
0.213
Nightly egg temperature
21.09
2.07
21.57
3.52
−0.52
0.608
Ambient mean temperature
23.56
1.23
23.33
2.04
0.33
0.743
Day egg temperature var
5.00
1.67
3.21
2.88
2.39
0.024*
Nightly egg temperature var
0.39
0.23
0.47
0.54
−0.45
0.669
Average incubation duration
75.70
29.40
38.41
43.56
−3.06
0.002**
Number of bouts
13.01
12.87
31.19
11.07
−4.30
< 0.001***
Total incubation duration
710.97
119.41
710.54
133.78
0.01
0.993
Nest attentiveness
49.41
8.31
52.03
8.78
−0.82
0.415
Offset
0.14
0.02
0.14
0.02
−0.86
0.437
Onset
0.90
0.03
0.84
0.07
2.77
0.009**
Significant levels of p values at 0.05, 0.01 and 0.001 were shown as *, ** and ***.
Figure
2.
Female Chinese Blackbird incubation behaviors over the 18 days of incubation, including average incubation duration (A), number of bouts (B), average off-bouts duration (C), and nest attentiveness (D). Urban site and females are in red, dotted lines, and rural site and females are in blue. Shown are means ± SE. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
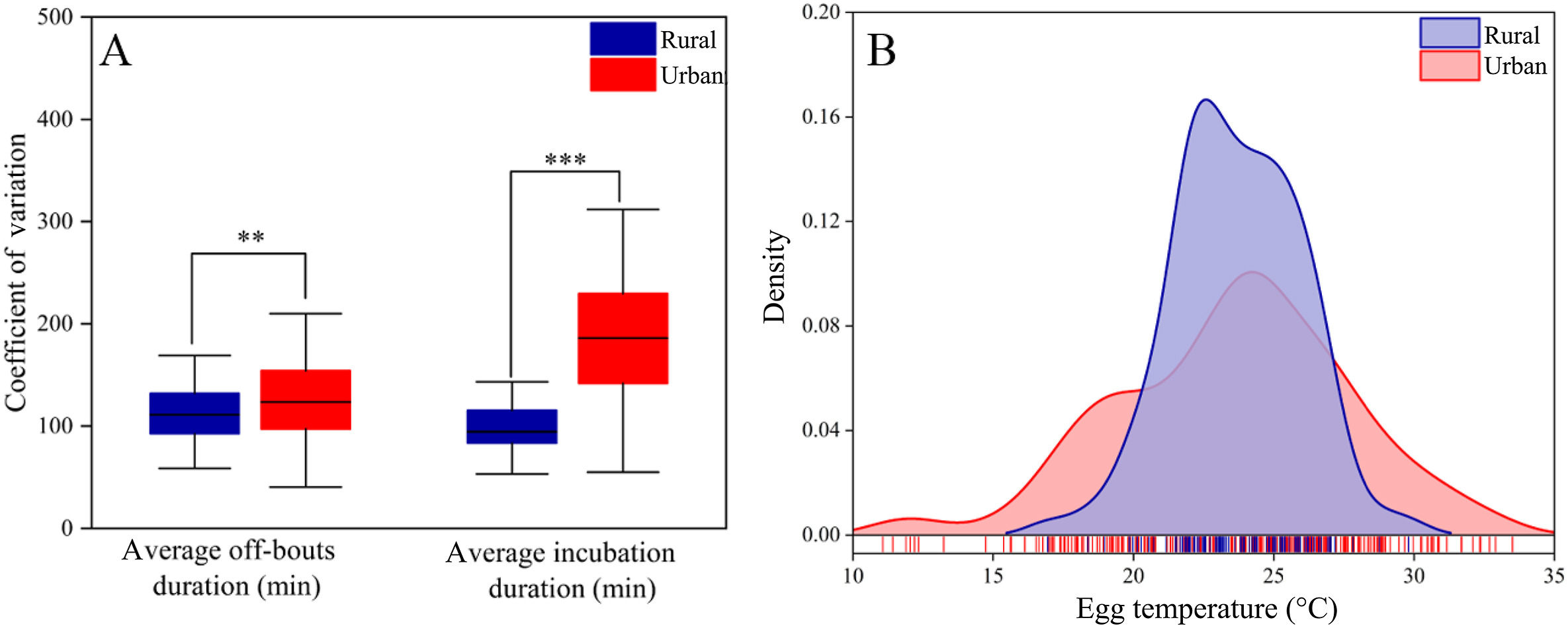
The coefficients of variation for day off-bout were respectively 125.91 ± 44.57 in urban areas and 113.22 ± 43.12 in rural areas, while the coefficients of variation for day incubation time were separately 182.72 ± 59.26 in urban areas and 102.96 ± 37.63 in rural areas. We also found that both urban female coefficients of variation for incubation and resting time were significantly higher than those of rural females, respectively (average off-bouts duration: t = −2.60, df = 437, p = 0.009; average incubation duration: t = −13.19, df = 437, p < 0.001; Fig. 3A). In addition, kernel density estimation curves for urban and rural Chinese Blackbird egg temperatures indicated a greater range of temperature variation in urban egg, while a smaller range in rural areas (Fig. 3B). However, the egg temperature distribution was concentrated around 15–30 ℃ at both sites.
Figure
3.
Variation in incubation behavior and temperature in urban and rural areas. (A) Coefficient of variation in the average off-bouts duration and the average incubation duration for female Chinese Blackbirds in urban and rural habitats. Whiskers show standard error, significance levels are indicated by asterisks (*p = 0.05–0.01, **p = 0.01–0.001, ***p < 0.001). (B) Kernel density estimation curve of egg daytime temperature for Chinese Blackbirds in urban and rural habitats.
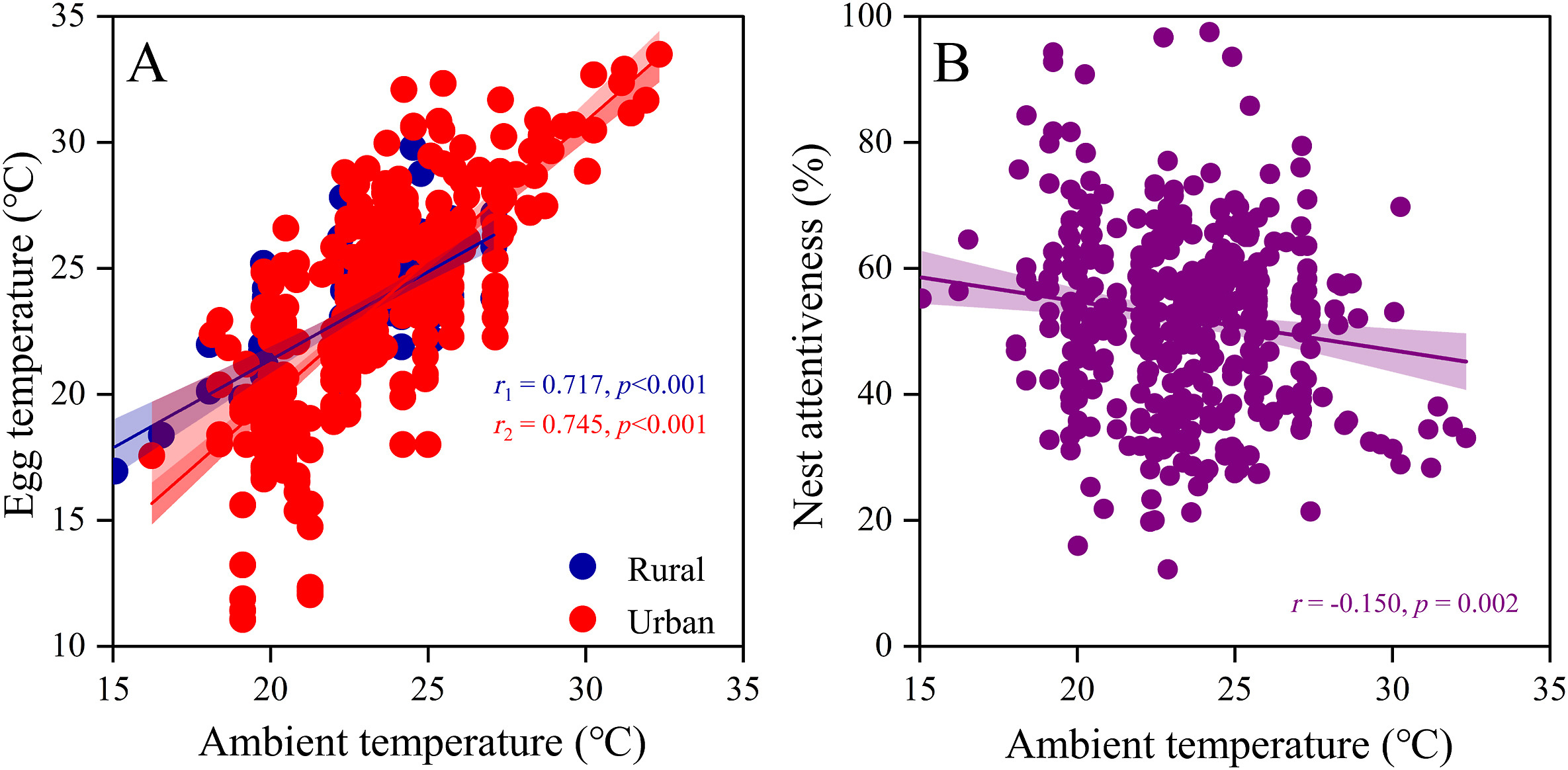
Significant positive correlations between egg temperature and ambient mean temperature in urban (r = 0.75, df = 437, p < 0.001) and rural (r = 0.72, df = 437, p < 0.001) areas (Fig. 4A), and their regression coefficients are significantly different (t = −2.84, df = 437, p = 0.005) from each other. However, the nest attentiveness of Chinese Blackbirds decreased significantly with increasing ambient mean temperature (r = −0.15, df = 437, p = 0.002; Fig. 4B). In urban areas, the average incubation duration of female was positively correlated (r = 0.13, df = 437, p = 0.018) with Julian date, while in rural areas it was negatively correlated (r = −0.27, df = 437, p = 0.005). The two regression lines have opposite trends of the two regions (Appendix Fig. S1), and their regression coefficients are significantly different (t = 2.23, df = 437, p = 0.027) from each other.
Figure
4.
Ambient mean temperature is strongly associated with (A) egg daytime temperature in urban and rural areas and (B) nest attentiveness in the female Chinese Blackbirds. Urban site and females are in red points, and rural site and females are in blue points. Linear regressions with 95% confidence intervals are shown within each panel, and lines represent the best fit from a linear model. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
3.3
Variation in urban and rural bird incubation behavior
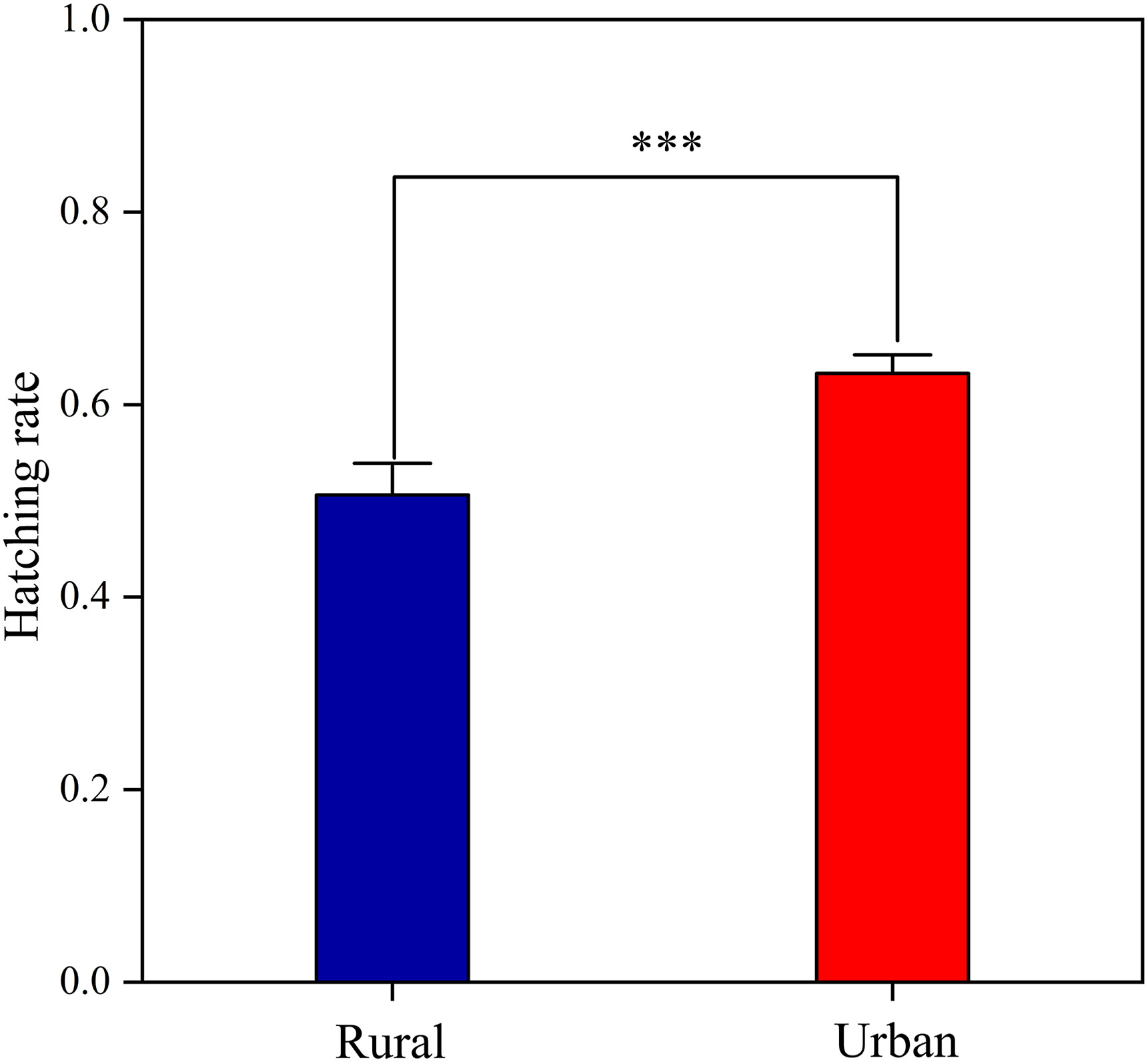
When comparing different incubation parameters of the Chinese Blackbird beween two regions, the independent samples t-test showed a significant difference among the four incubation parameters of Chinese Blackbirds (Table 1). The variation of egg temperature in urban areas (3.21 ± 2.88) was significantly less than that in rural areas (5.00 ± 1.67) at the level of 0.05, and the same is true for the average incubation duration and the latest time to on-bouts (Table 1). Furthermore, the hatching rate (urban: 0.53 ± 0.41; rural: 0.40 ± 0.01) was also significantly higher in urban females than in rural females (Fig. 5).
Figure
5.
Hatching rate of female Chinese Blackbirds in urban and rural habitats. Whiskers show standard error, and significance levels are indicated by asterisks (***p < 0.001).
The binomial generalized linear model results also demonstrate similar results (Table 2). We identified one of the best models (ΔAIC = 0.000) for urban and rural. The incubation factors include day incubation temperature var, number of bouts, and onset. The results of logistic regression indicated that the model (wi = 0.99; day incubation temperature + the number of bouts + onset) was the top-ranked model for predicting the critical incubation factors of the urban and rural areas, and the predictive capability of the model was excellent (R2 = 0.45). All parameters in this model had significant effects (Table 2).
Table
2.
Model-averaged summary statistics of best fitting GLM of Chinese Blackbird incubation parameters variation in urban and rural areas.
Empty Cell
Estimate
SE
t value
p value
Intercept
4.41
2.36
1.87
0.062
Day incubation temperature var
−0.07
0.03
−2.80
0.005**
Number of bouts
0.09
0.01
7.67
< 0.001***
Onset
−5.30
2.61
−2.03
0.042*
Significant levels of p values at 0.05, 0.01 and 0.001 were shown as *, ** and ***.
Key incubation factors affecting the DSR of Chinese Blackbirds in urban and rural areas were analyzed using a negative binomial generalized linear model (Table 3). We identified one of the best models of DSR among the two regions, respectively. The factors, including day incubation temperature and the number of bouts, appeared in two models. This relationship was positive for two regions (day survival rate increases with increasing day incubation temperature). Nevertheless, the number of bouts of urban females had a significantly positive effect on DSR while the DSR of rural females decreased with an increasing number of bouts (Table 3).
Table
3.
Model-averaged summary statistics of best fitting GLM of Chinese Blackbird incubation parameters variation on day survival rate in urban and rural areas.
Day survival rate
Variable
Estimate
SE
t value
p value
Urban
Intercept
−0.14
0.22
−0.66
0.507
Day incubation temperature
0.01
0.01
2.21
0.028*
Day incubation temperature var
−0.01
0.01
−2.11
0.036*
Nightly incubation temperature var
−0.13
0.03
−3.94
< 0.001***
Number of bouts
0.00
0.00
2.90
0.004**
Onset
−0.92
0.21
−4.32
< 0.001***
Rural
Intercept
−1.86
0.31
−6.09
< 0.001***
Day incubation temperature
0.04
0.01
3.49
< 0.001***
Number of bouts
−0.00
0.00
−2.31
0.023*
Significant levels of p values at 0.05, 0.01 and 0.001 were shown as *, ** and ***.
As demonstrated by the results of negative binomial regression, the model (wi = 0.57; day incubation temperature + day incubation temperature var + nightly incubation temperature var + number of bouts + onset) was the best model for predicting nest DSR by the urban juvenile, and it had excellent predictive accuracy (R2 = 0.45). The best model (wi = 0.52) of rural females only contained two variables (day incubation temperature + number of bouts), and the predictive capability of the model was acceptable (R2 = 0.45). All parameters in the model had significant effects at the 0.05 level (Table 3).
The results were in the opposite direction of our predictions. Higher fitness of urban individuals under lower environmental pressure is not achieved by decreasing the quantity of energy invested in incubation. Our survey indicated that total incubation duration and nest attentiveness have no difference between urban and rural areas over the entire incubation period (Table 1; Fig. 2). Urban females had more and shorter day incubation bouts (Fig. 2) and a higher number of bouts per day, which means that females were on and off the eggs more frequently compared to rural females (Table 1). Bird must cope with varying environmental conditions and adjust the balance between incubation and self-maintenance (Nord and Cooper, 2020; Seress et al., 2020; Ton et al., 2021). Urban areas often experience higher disturbance from traffic noise (Cronin et al., 2022), night light interference (Nordt and Klenke, 2013), and human disturbance (Lovell et al., 2022; Osugi et al., 2022), which all may force urban females to leave the nest more frequently. Furthermore, our study also suggested that incubation temperature is a crucial factor affecting the DSR rate in two regions (Table 3). Species of animals and plants have evolved to adapt to and acclimate to the temperature and microhabitat conditions. Urban warmer ambient temperature (Fig. 1), combined with previously described lower predation risk and food availability (Conway and Martin, 2000), may drive selective forces in urban birds. This selection pressure causes birds to adjust their incubation behavior (select more frequent off-bouts at consistent total incubation duration and nest attentiveness) to achieve higher fitness.
Temperature is one of the most critical aspects of successful incubation. Ambient temperature is among the strongest drivers of avian incubation behavior (Conway and Martin, 2000; Ardia et al., 2009). The urban heat island, a byproduct of urbanization in which cities have higher temperatures than surrounding natural lands, could have lasting effects (Hall and Warner, 2019). We found that urban sites have warmer micro-environmental temperatures (Fig. 1) and as the ambient temperature increases, Chinese Blackbirds become less attentive to their nests (Fig. 2). Adults respond to elevated temperatures by decreasing incubation effort due to reducing thermostatic demand (Ton et al., 2021). Higher temperatures contribute to more stable conditions, with warmer nest temperatures. The egg cooling rate is slower, and egg temperature does not drop under higher ambient temperature (Szentirmai et al., 2005; McClintock et al., 2014). Therefore, urban females could use this extra time to acquire more energy or spend more time on self-maintenance activities (Londono et al., 2008), which contributed to incubation success.
One of the key factors influencing incubation behavior is predation which leads directly to reproductive failure in birds (Abbey-Lee and Dingemanse, 2019; Chen et al., 2022). Following our expectations, rural birds face a larger risk of predation, and rural females take fewer but longer off-bouts from the nests (Table 2). These results support the original hypothesis that nest predation risk is a critical factor in parental activity at the nest described by Conway and Martin (2000). This behavior could reduce nest predation during incubation in the rural areas, but it may also occur at the cost of increasing exposure of eggs to deleterious temperatures (Thompson et al., 2022). Accordingly, urban birds may move on and off the nest more frequently with shorter bouts (Table 1), which could alleviate predation risk in urbanized habitats. Accordingly, this strategy might be feasible for urban birds because such birds have access to spatially and temporally predictable ad libitum sources of high-quality anthropogenic food (Schoech and Bowman, 2003), which permits urban females to forage higher quality food than rural females (Chamberlain et al., 2018). Short, frequent off-bouts may be an ideal strategy for optimizing fitness while minimizing nest predators.
During the incubation period, food must be provided promptly so that the energy consumed dramatically may be supplemented sufficiently. The high proportion of impermeable surface area decreases invertebrates (including worms and insects) in the urban landscape. Because invertebrates, such as earthworms, are the primary food source for blackbirds during the breeding season. Food availability for urban blackbirds to receive the energy needed for hatching is limited (Charmantier et al., 2017). Food shortages might exacerbate the frequency of off-bouts in urban blackbirds.
4.2
Higher plasticity facilitates the successful settlement of birds in cities
Individuals who successfully colonize in urban environments have higher behavioral plasticity in incubation strategies. In this study, we provided evidence for the complex incubation behavioral plasticity of the Chinese Blackbird that likely enhanced the successful colonization of the urbanized landscape. Compared with rural females, the variation range of incubation behavior and temperature of urban individuals was higher (Fig. 3A), meaning a wider niche. Further, the number of bouts was found to positively affect the survival rate of urban birds' nests while negatively affecting the rate of survival for rural birds (Table 3). Hence, behavioral plasticity may be beneficial for females in urban environments.
Adjusting incubation behavior can help birds adapt to the current environmental difficulties, ensure an optimal thermal environment for the developing embryos, and allow birds to inhabit urbanized areas (Minias and Tregenza, 2015; Heppner and Ouyang, 2021). Even when eggs are left unattended, warmer ambient temperatures and lower predation pressure allow urban birds to pay less attention to nests (Lundblad and Conway, 2021a, b). Reduced energy expenditure during incubation and increased foraging time during the incubation period may positively affect parental condition, which in turn enables them to forage more effectively later while feeding nestlings, leading to improved nestling body condition (Ardia et al., 2010; Merrill et al., 2020; Ton et al., 2021). In comparison to rural birds, urban birds have a higher success rate in hatching offspring (Fig. 5). Moreover, the range of egg thermal gradients in urban blackbirds is more comprehensive than in rural regions (Fig. 3B). This physiological adaptation might be one of the reasons why urban blackbirds can have frequent off-bouts. Unfortunately, our study design did not allow us to assess separately the thermal tolerance of each egg, which may directly lead to embryo death. However, the vital role of phenotypic and physiological plasticity in colonization processes has recently been supported by the observation that the current rate of environmental change often exceeds the rate of evolutionary response in many populations (Bell and Collins, 2008; Chevin et al., 2010). This physiological adaptation may be a transitional stage in the evolutionary process (Swallow et al., 2005). Moreover, an urban region in our study has been urbanized in the recent 20 years (Han et al., 2019). Plastic behavioral responses may give additional time for genetic changes to arise and prevent populations from extinction in rapidly changing environments (Tuomainen and Candolin, 2011; Sol et al., 2012).
The process of urban colonization by birds often requires substantial alterations in the nest-site selection pattern. The availability of typical nest-site and micro-habitats may be limited in urban landscapes (Davis et al., 2014). Urban Chinese Blackbirds use different urban buildings as nesting substrates (Han et al., 2019), and it is worth noting that in our other studies on artificial nest boxes, urban Chinese Blackbirds also built their nests on top of the nest box (Appendix Fig. S2). Birds that have adapted to and actively use such nest sites usually have higher reproductive success, leading to a gradual increase in population size and range, and thus can settle successfully in urban areas (De Laet and Summers-Smith, 2007). In addition, anthropogenic disturbance reduces the risk of predation (St Clair et al., 2010), which means that birds nesting on buildings in human settlements tend to have weaker anti-predatory behavior (Møller and Diaz, 2018) and thus have more energy for incubation and brood production. Furthermore, decreased caterpillar availability in urban environments may constrain the female during egg-laying, limiting the number of eggs she can produce due to nutrient/calcium limitations (Caizergues et al., 2018). Compared to rural blackbirds in this study, urban individuals lay less clutch size (4.93 ± 0.85 and 5.21 ± 0.74 in urban and rural, respectively (unpublished data)), which supported limited clutch size in the urban population.
Although urbanization poses a suite of new challenges which influence lifetime fitness (Sepp et al., 2018), the incubation behavior, with the high plasticity of urban females, may be a critical factor in their ability to settle in urban landscapes successfully. Like most others in behavioral urban ecology (Heppner and Ouyang, 2021), our study used a single urban and rural site. Therefore, these results need to be replicated in the future in more cities as population differences have not always been generalized across urban and rural replicates. However, this study gives an initial insight into altered behaviors due to environmental change. By studying incubation behavior differences between urban and rural wildlife, we can better understand urbanization's effects on wildlife behavior and how these parental behaviors affect offspring phenotypes in a city landscape.
Authors' contributions
LM designed the experiments, analyzed the data, wrote the manuscript, and reviewed the drafts. YL, WL, Zheng Z, WL, Ziwei Z, XZ, CZ, JB and YH participated in field investigation and data collection. LR conceived the experiments, provided funding acquisition and supervision. All authors read and approved the final manuscript.
Ethics statement
All procedures applied during the research were in accordance with the guidelines for animal care outlined by Chinese wildlife conservation laws. We have all the required permissions for monitoring birds and measuring their habitats from Jiangxi Provincial Department of Forestry, China.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We gratefully acknowledge the contribution of Xinqi Lu, Mingwang Chen, Xi Zhu and Guanglong Sun for the fieldwork. The study was supported by financial support from the Key Basic Science and Technology Project of the Ministry of Science and Technology, China under Grant (2015FY110200) and Jiangxi Provincial Natural Science Foundation, China (No. 20224ACB205006).
Abbey-Lee, R.N., Dingemanse, N.J., 2019. Adaptive individual variation in phenological responses to perceived predation levels. Nat. Commun. 10, 1601.
Ardia, D.R., Perez, J.H., Chad, E.K., Voss, M.A., Clotfelter, E.D., 2009. Temperature and life history: experimental heating leads female tree swallows to modulate egg temperature and incubation behaviour. J. Anim. Ecol. 78, 4-13.
Ardia, D.R., Perez, J.H., Clotfelter, E.D., 2010. Experimental cooling during incubation leads to reduced innate immunity and body condition in nestling tree swallows. Proc. R. Soc. B-Biol. Sci. 277, 1881-1888.
Bailey, L.D., van de Pol, M., Adriaensen, F., Arct, A., Barba, E., Bellamy, P.E., et al., 2022. Bird populations most exposed to climate change are less sensitive to climatic variation. Nat. Commun. 13, 2112.
Bailly, J., Scheifler, R., Berthe, S., Clement-Demange, V.A., Leblond, M., Pasteur, B., et al., 2016. From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 157, 377-392.
Batisteli, A.F., De Souza, L.B., Santieff, I.Z., Gomes, G., Soares, T.P., Pini, M., et al., 2021. Buildings promote higher incubation temperatures and reduce nest attentiveness in a Neotropical thrush. Ibis 163, 79-89.
Batisteli, A.F., Pizo, M.A., Sarmento, H., 2022. Female neophobia predicts the use of buildings as nesting sites in a Neotropical songbird. Anim. Behav. 183, 151-157.
Bell, G., Collins, S., 2008. Adaptation, extinction and global change. Evol. Appl. 1, 3-16.
Bujan, J., Charavel, E., Bates, O.K., Gippet, J.M.W., Darras, H., Lebas, C., et al., 2021. Increased acclimation ability accompanies a thermal niche shift of a recent invasion. J. Anim. Ecol. 90, 483-491.
Caizergues, A.E., Gregoire, A., Charmantier, A., 2018. Urban versus forest ecotypes are not explained by divergent reproductive selection. Proc. R. Soc. B-Biol. Sci. 285, e20180261.
Capilla-Lasheras, P., 2018. incR: a new R package to analyse incubation behaviour. J. Avian Biol. 49, e01710.
Carvalho, A.V., Hennequet-Antier, C., Crochet, S., Bordeau, T., Courousse, N., Cailleau-Audouin, E., et al., 2020. Embryonic thermal manipulation has short and long-term effects on the development and the physiology of the Japanese quail. PLoS ONE 15, e0227700.
Chamberlain, M.J., Wightman, P.H., Cohen, B.S., Collier, B.A., 2018. Gobbling activity of eastern wild turkeys relative to male movements and female nesting phenology in South Carolina. Wildlife Soc. Bull. 42, 632-642.
Charmantier, A., Demeyrier, V., Lambrechts, M., Perret, S., Gregoire, A., 2017. Urbanization is associated with divergence in pace-of-life in Great Tits. Front. Ecol. Evol. 5, 53.
Chen, X., Zhang, Q., Lan, S., Chen, S., Wang, Y., 2022. Nest predation pressure on Chinese Bulbuls decreases along the urbanization gradient in Hangzhou, China. Avian Res. 13, 100049.
Chevin, L.M., Lande, R., Mace, G.M., 2010. Adaptation, plasticity, and extinction in a changing environment: towards a predictive theory. PLoS Biol. 8, e1000357.
Conway, C.J., Martin, T.E., 2000. Evolution of passerine incubation behavior: influence of food, temperature, and nest predation. Evolution 54, 670-685.
Corsini, M., Schöll, E.M., Di Lecce, I., Chatelain, M., Dubiec, A., Szulkin, M., 2021. Growing in the city: urban evolutionary ecology of avian growth rates. Evol. Appl. 14, 69-84.
Costa, B.T.A., Lopes, T.S.B., Mesquita, M.A., Lara, L.J.C., Araujo, I.C.S., 2020. Thermal manipulations of birds during embryogenesis. World. Poultry Sci. J. 76, 843-851.
Crates, R.A., Firth, J.A., Farine, D.R., Garroway, C.J., Kidd, L.R., Aplin, L.M., et al., 2016. Individual variation in winter supplementary food consumption and its consequences for reproduction in wild birds. J. Avian Biol. 47, 678-689.
Cronin, A.D., Smit, J.A.H., Munoz, M.I., Poirier, A., Moran, P.A., Jerem, P., et al., 2022. A comprehensive overview of the effects of urbanisation on sexual selection and sexual traits. Biol. Rev. 97, 1325-1345.
Croston, R., Hartman, C.A., Herzog, M.P., Casazza, M.L., Feldheim, C.L., Ackerman, J.T., 2020. Timing, frequency, and duration of incubation recesses in dabbling ducks. Ecol. Evol. 10, 2513-2529.
Curtis, J.R., Robinson, W.D., Rompré, G., Austin, S.H., 2022. Urbanization is associated with unique community simplification among birds in a neotropical landscape. Landscape Ecol. 37, 209-231.
Davis, A., Major, R.E., Taylor, C.E., 2014. Distribution of tree-hollows and hollow preferences by parrots in an urban landscape. Emu 114, 295-303.
Dawson, A., King, V.M., Bentley, G.E., Ball, G.F., 2001. Photoperiodic control of seasonality in birds. J. Biol. Rhythm. 16, 365-380.
De Laet, J., Summers-Smith, J.D., 2007. The status of the urban house sparrow Passer domesticus in north-western Europe: a review. J. Ornithol. 148, 275-278.
Deeming, D.C., 2002. Avian Incubation: Behaviour, Environment and Evolution. Oxford University Press, Oxford.
Deeming, D.C., Gray, L.A., 2016. Incubation attentiveness and nest insulatory values correlate in songbirds. Avian Biol. Res. 9, 32-36.
Fenoglio, M.S., Calvino, A., Gonzalez, E., Salvo, A., Videla, M., 2021. Urbanisation drivers and underlying mechanisms of terrestrial insect diversity loss in cities. Ecol. Entomol. 46, 757-771.
Fontaine, J.J., Martin, T.E., 2006. Parent birds assess nest predation risk and adjust their reproductive strategies. Ecol. Lett. 9, 428-434.
González-Lagos, C., Cardador, L., Sol, D., 2021. Invasion success and tolerance to urbanization in birds. Ecography 44, 1642-1652.
Grimaudo, A.T., Hope, S.F., DuRant, S.E., Kennamer, R.A., Hallagan, J.J., Hopkins, W.A., 2020. Ambient temperature and female body condition are related to night incubation behavior in wood ducks Aix sponsa. J. Avian Biol. 51, e02379.
Grimm, N.B., Faeth, S.H., Golubiewski, N.E., Redman, C.L., Wu, J., Bai, X., et al., 2008. Global change and the ecology of cities. Science 319, 756-760.
Hall, J.M., Warner, D.A., 2019. Thermal tolerance in the urban heat island: thermal sensitivity varies ontogenetically and differs between embryos of two sympatric ectotherms. J. Exp. Biol. 222, jeb210708.
Han, Y.Q., Bai, J.P., Zhang, Z., Wu, T., Chen, P., Sun, G.L., et al., 2019. Nest site selection for five common birds and their coexistence in an urban habitat. Sci. Total Environ. 690, 748-759.
Havlíček, J., Riegert, J., Bandhauerová, J., Fuchs, R., Šálek, M., 2021. Species-specific breeding habitat association of declining farmland birds within urban environments: conservation implications. Urban Ecosyst. 24, 1259-1270.
Heppner, J.J., Ouyang, J.Q., 2021. Incubation behavior differences in urban and rural House Wrens, Troglodytes aedon. Front. Ecol. Evol. 9, 590069.
Hope, S.F., Hopkins, W.A., Angelier, F., 2021. Parenting in the city: does urbanization influence avian incubation behavior? Integr. Comp. Biol. 61, 389-390.
Kettel, E.F., Gentle, L.K., Quinn, J.L., Yarnell, R.W., 2018. The breeding performance of raptors in urban landscapes: a review and meta-analysis. J. Ornithol. 159, 1-18.
Kleindorfer, S., Common, L.K., O'Connor, J.A., Garcia-Loor, J., Katsis, A.C., Dudaniec, R.Y., et al., 2021. Female in-nest attendance predicts the number of ectoparasites in Darwin's finch species. Proc. R. Soc. B-Biol. Sci. 288, e20211668.
Kurucz, K., Purger, J.J., Batáry, P., 2021. Urbanization shapes bird communities and nest survival, but not their food quantity. Global Ecol. Conserv. 26, e01475.
Lilliefors, H., 1987. Auld simulation results and when to stop. Am. Stat. 41, 243-244.
Londono, G.A., Levey, D.J., Robinson, S.K., 2008. Effects of temperature and food on incubation behaviour of the northern mockingbird, Mimus polyglottos. Anim. Behav. 76, 669-677.
Lovell, C., Li, S.Y., Turner, J., Carbone, C., 2022. The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: the results of an intensive camera trap study. Ecol. Evol. 12, e8746.
Lundblad, C.G., Conway, C.J., 2021a. Intraspecific variation in incubation behaviours along a latitudinal gradient is driven by nest microclimate and selection on neonate quality. Funct. Ecol. 35, 1028-1040.
Lundblad, C.G., Conway, C.J., 2021b. Nest microclimate and limits to egg viability explain avian life-history variation across latitudinal gradients. Ecology 102, e03338.
Luscier, J.D., 2022. A core on the Atlantic margin of Europe: an urban bird assemblage in Cork City, Ireland. Urban Ecosyst. 25, 585-599.
McClintock, M.E., Hepp, G.R., Kennamer, R.A., 2014. Plasticity of incubation behaviors helps Wood Ducks (Aix sponsa) maintain an optimal thermal environment for developing embryos. Auk 131, 672-680.
McIntyre, N.E., Rango, J., Fagan, W.F., Faeth, S.H., 2001. Ground arthropod community structure in a heterogeneous urban environment. Landscape Urban Plann. 52, 257-274.
Merrill, L., Ospina, E.A., Santymire, R.M., Benson, T.J., 2020. Egg incubation temperature affects development of innate immune function in nestling American Robins (Turdus migratorius). Physiol. Biochem. Zool. 93, 1.
Minias, P., Tregenza, T., 2015. Successful colonization of a novel urban environment is associated with an urban behavioural syndrome in a reed-nesting waterbird. Ethology 121, 1178-1190.
Møller, A.P., Diaz, M., 2018. Avian preference for close proximity to human habitation and its ecological consequences. Curr. Zool. 64, 623-630.
Nord, A., Cooper, C.B., 2020. Night conditions affect morning incubation behaviour differently across a latitudinal gradient. Ibis 162, 827-835.
Nord, A., Nilsson, J.A., 2011. Incubation temperature affects growth and energy metabolism in Blue Tit nestlings. Am. Nat. 178, 639-651.
Nord, A., Williams, J.B., 2015. The energetic costs of incubation. In: Deeming, D.C., Reynolds, S.J. (Eds.), Nests, Eggs, and Incubation. Oxford University Press, Oxford, pp. 152–170.
Nordt, A., Klenke, R., 2013. Sleepless in town-drivers of the temporal shift in dawn song in urban European blackbirds. PLoS ONE 8, e71476.
Osugi, S., Trentin, B.E., Koike, S., 2022. Effects of human activity on the fallen-fruit foraging behavior of carnivoran species in an urban forest. Mammal Study 47, 113-123.
Plaza, M., Cantarero, A., Cuervo, J.J., Moreno, J., 2018. Female incubation attendance and nest vigilance reflect social signaling capacity: a field experiment. Behav. Ecol. Sociobiol. 72, 24.
Ryeland, J., House, C.M., Umbers, K.D.L., Spencer, R.J., 2021. Optimal clutch size and male incubation investment in the male-only incubating emu (Dromaius novaehollandiae). Behav. Ecol. Sociobiol. 75, 168.
Schoech, S.J., Bowman, R., 2003. Does differential access to protein influence differences in timing of breeding of Florida scrub-jays (Aphelocoma coerulescens) in suburban and wildland habitats? Auk 120, 1114-1127.
Sepp, T., McGraw, K.J., Kaasik, A., Giraudeau, M., 2018. A review of urban impacts on avian life-history evolution: does city living lead to slower pace of life? Global Change Biol. 24, 1452-1469.
Seress, G., Sandor, K., Evans, K.L., Liker, A., 2020. Food availability limits avian reproduction in the city: an experimental study on great tits Parus major. J. Anim. Ecol. 89, 1570-1580.
Smith, J.A., Gaynor, K.M., Suraci, J.P., 2021. Mismatch between risk and response may amplify lethal and non-lethal effects of humans on wild animal populations. Front. Ecol. Evol. 9, 604973.
Sol, D., Bartomeus, I., Griffin, A.S., 2012. The paradox of invasion in birds: competitive superiority or ecological opportunism? Oecologia 169, 553-564.
St Clair, J.J.H., Garcia-Pena, G.E., Woods, R.W., Szekely, T., 2010. Presence of mammalian predators decreases tolerance to human disturbance in a breeding shorebird. Behav. Ecol. 21, 1285-1292.
Swallow, J.G., Rhodes, J.S., Garland, T., 2005. Phenotypic and evolutionary plasticity of organ masses in response to voluntary exercise in house mice. Integr. Comp. Biol. 45, 426-437.
Szentirmai, I., Szekely, T., Liker, A., 2005. The influence of nest size on heat loss of Penduline Tit eggs. Acta Zool. Acad. Sci. H. 51, 59-66.
Thompson, M.J., Capilla-Lasheras, P., Dominoni, D.M., Reale, D., Charmantier, A., 2022. Phenotypic variation in urban environments: mechanisms and implications. Trends Ecol. Evol. 37, 171-182.
Thomson, D.L., Monaghan, P., Furness, R.W., 1998. The demands of incubation and avian clutch size. Biol. Rev. 73, 293-304.
Ton, R., Hurley, L.L., Griffith, S.C., 2021. Higher experimental ambient temperature decreases female incubation attentiveness in Zebra Finches (Taeniopygia guttata) and lower effort yields negligible energy savings. Ibis 163, 1045-1055.
Vincze, E., Bókony, V., Garamszegi, L.Z., Seress, G., Pipoly, I., Sinkovics, C., et al., 2021. Consistency and plasticity of risk-taking behaviour towards humans at the nest in urban and forest great tits, Parus major. Anim. Behav. 179, 161-172.
Wang, Y.P., Chen, S.H., Blair, R.B., Jiang, P.P., Ding, P., 2009. Nest composition adjustments by Chinese Bulbuls Pycnonotus sinensis in an urbanized landscape of Hangzhou (E China). Acta Ornithol. 44, 185-192.
Webb, G.J.W., Cooperpreston, H., 1989. Effects of incubation-temperature on crocodiles and the evolution of reptilian oviparity. Am. Zool. 29, 953-971.
Yoon, J., Jung, J.S., Joo, E.J., Kim, B.S., Park, S.R., 2017. Parent birds assess nest predation risk: influence of cavity condition and avian nest predator activity. J. Avian Biol. 48, 691-699.
Zhang, Y.M., Zhao, D.Q., Ruan, L.Z., Dong, Y.H., Wang, H., Fasola, M., 2003. Paring age and reproductive success in black-crowned night herons at Tai Lake, China. Zool. Res. 1, 54-56.
Zhou, B., Liu, J., Liang, W., 2020. Breeding in a noisy world: attraction to urban arterial roads and preference for nest-sites by the scaly-breasted munia (Lonchura punctulata). Global Ecol. Conserv. 22, e00987.
Zhu, C., Li, W., Gregory, T., Wang, D., Ren, P., Zeng, D., et al., 2021. Arboreal camera trapping: a reliable tool to monitor plant-frugivore interactions in the trees on large scales. Remote Sens. Ecol. Conserv. 8, 92-104.
Table
3.
Model-averaged summary statistics of best fitting GLM of Chinese Blackbird incubation parameters variation on day survival rate in urban and rural areas.
Day survival rate
Variable
Estimate
SE
t value
p value
Urban
Intercept
−0.14
0.22
−0.66
0.507
Day incubation temperature
0.01
0.01
2.21
0.028*
Day incubation temperature var
−0.01
0.01
−2.11
0.036*
Nightly incubation temperature var
−0.13
0.03
−3.94
< 0.001***
Number of bouts
0.00
0.00
2.90
0.004**
Onset
−0.92
0.21
−4.32
< 0.001***
Rural
Intercept
−1.86
0.31
−6.09
< 0.001***
Day incubation temperature
0.04
0.01
3.49
< 0.001***
Number of bouts
−0.00
0.00
−2.31
0.023*
Significant levels of p values at 0.05, 0.01 and 0.001 were shown as *, ** and ***.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds