Cristel Álvarez-Castillo, Ian MacGregor-Fors, Stefan L. Arriaga-Weiss, Claudio Mota-Vargas, Diego Santiago-Alarcon. 2022: Abundance of White-fronted Parrots and diet of an urban parrot assemblage (Aves: Psittaciformes) in a green Neotropical city. Avian Research, 13(1): 100019. DOI: 10.1016/j.avrs.2022.100019
Citation:
Cristel Álvarez-Castillo, Ian MacGregor-Fors, Stefan L. Arriaga-Weiss, Claudio Mota-Vargas, Diego Santiago-Alarcon. 2022: Abundance of White-fronted Parrots and diet of an urban parrot assemblage (Aves: Psittaciformes) in a green Neotropical city. Avian Research, 13(1): 100019. DOI: 10.1016/j.avrs.2022.100019
Cristel Álvarez-Castillo, Ian MacGregor-Fors, Stefan L. Arriaga-Weiss, Claudio Mota-Vargas, Diego Santiago-Alarcon. 2022: Abundance of White-fronted Parrots and diet of an urban parrot assemblage (Aves: Psittaciformes) in a green Neotropical city. Avian Research, 13(1): 100019. DOI: 10.1016/j.avrs.2022.100019
Citation:
Cristel Álvarez-Castillo, Ian MacGregor-Fors, Stefan L. Arriaga-Weiss, Claudio Mota-Vargas, Diego Santiago-Alarcon. 2022: Abundance of White-fronted Parrots and diet of an urban parrot assemblage (Aves: Psittaciformes) in a green Neotropical city. Avian Research, 13(1): 100019. DOI: 10.1016/j.avrs.2022.100019
Urban ecosystems are evolutionarily recent novel environments acting as biodiversity filters. Psittacidae birds are considered successful urban adapters mainly due to their generalist feeding and opportunistic behavior, allowing them to occupy environments from cold temperate to dry xeric areas. Therefore, it is important to understand how these species interact in the urban environment. We studied the interannual (2013–2016) abundance of the White-fronted Parrot (Amazona albifrons) in the Neotropical cities of Xalapa and Coatepec, in Central Veracruz, México. Additionally, we studied the feeding ecology during 13 months of 6 parrot species detected in the city of Xalapa. The abundance of the White-fronted Parrot was significantly higher in Xalapa than in Coatepec, and it was homogeneous across years. Non-native plants represented 30–41% of Psittacidae diets in Xalapa, where seeds were the most commonly consumed resource. We recorded the highest Psittacidae species richness and highest diet overlap among species by the end of the dry season (April–May). The White-fronted Parrot had the highest plant richness in its diet, followed by the Monk Parakeet (Myiopsitta monachus) and the Green Parakeet (Psittacara holochlorus); yet, the White-fronted Parrot had a specialized diet dominated by two plant species (Grevillea robusta and Ficus aurea). The diet overlap among the three above-mentioned parrot species was not significantly different to a null model, where the White-fronted Parrot and the Monk Parakeet overlapped during the months of February, April, June, and September. The White-fronted Parrot is an urban adapter that has successfully expanded its geographic range via natural means and by human activities. The invasive Monk Parakeet is currently restricted to one park in Xalapa, and it has remained in that stage for many years (i.e., pre-expansion phase). Exotic plant species in Xalapa represent ∼55% of the woody vegetation, some of which have longer flowering and fruiting periods that may have aided the successful establishment of parrot species in urban environments.
Anthropogenic activities are a threat to biodiversity, in particular urban development and its associated urban metabolism that extends human impacts for hundreds of kilometers beyond city borders (Adler and Tanner, 2013; Maxwell et al., 2016). The drastic changes of environmental conditions entailed by urbanization act as a filter for species richness and phylogenetic diversity, imposing selective pressures forcing species to adapt or to completely avoid this novel habitat (Aronson et al., 2014; Sol et al., 2017). In this sense, Fischer et al. (2015) have classified species based on their population biology as urban dwellers, urban utilizers and urban avoiders. Dwellers are those species that conduct all their activities within the urban environment – in this group is where we encounter many non-native and native invasive species – and do not depend on natural environments to survive; utilizers are those that take advantage of some city resources (e.g., greenspaces, blue spaces) mostly to feed and rarely to breed, but that depend on non-urban environments for population viability; and avoiders are those species unable to adapt and survive within urban environments, but they may use some city resources such as parks (Fischer et al., 2015). The category under which species fall depends on factors such as diet, migratory behavior, reproductive behavior and if they are native or exotic (Kark et al., 2007; Shanahan et al., 2014). Therefore, organisms’ tolerance to environmental conditions depends on ecological, behavioral, and physiological flexibility (Bonier et al., 2007).
Within urban environments, greenspaces (e.g., parks, yards, natural preserves) are areas that provide unique resources across the year, which favors the presence of species (e.g., urban adapters) that need conditions similar to those found in natural environments, becoming biodiversity refuges within cities (Davis et al., 2012, 2015). In the specific case of birds, the order Psittaciformes includes many species that have taken advantage of greenspaces in cities across the world, including areas outside their natural distribution where they have escaped captivity (Menchetti and Mori, 2014), and where they can become invasive (e.g., Myiopsitta monachus; MacGregor-Fors et al., 2011). In a sense, cities’ greenspaces can act as refuges for species of parrots, which is important given their vulnerability caused by illegal trade (Berkunsky et al., 2017). Success of parrots within cities is believed to be due to a generalist diet that allows them to use abundant resources throughout the year by modifying their diet as a function of food availability, as well as their opportunistic behavior (i.e., rapid colonization and impeding the establishment of other species; Gascon et al., 1999; Renton, 2001; Muñoz and Real, 2006; Boyes and Perrin, 2009; Martens et al., 2013). Yet, the natural history of Psittaciformes in urban environments is poorly known, precluding us to understand how these species interact and use available resources across the year.
In this study, we aim to understand the abundance and feeding behavior of parrots within the Neotropical city of Xalapa in México. Particularly, we aim first to investigate how the abundance of the most common species – the White-fronted Parrot (Amazona albifrons) – varies across different years both in the city of Xalapa and in the nearby city of Coatepec, and second to determine the diet of different parrot species across a full year. Based on parrot species sizes, we first expected that birds of similar size had a higher overlap in their diets, and second that larger species had a higher diet breadth given that their stronger and larger beaks allow them to use resources such as large hard seeds that smaller species cannot crack. Finally, we expected a constant abundance of White-fronted Parrots across years given that they have a generalist diet, and as opportunistic species they are able to hoard and protect suitable territories across the year.
2.
Materials and methods
2.1
Study site
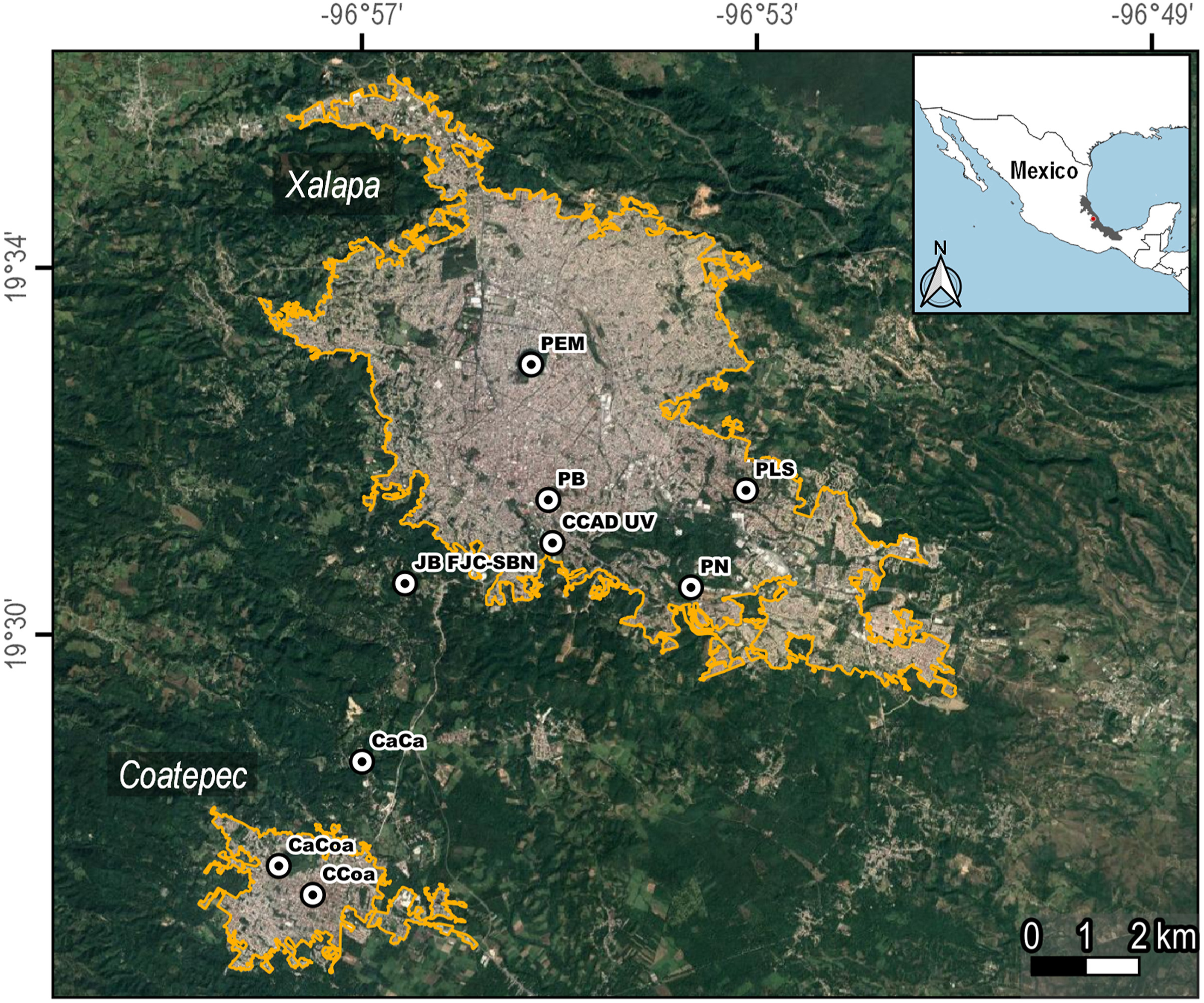
To compare White-fronted Parrot relative abundance between years, we conducted surveys and counted the number of White-fronted Parrots in Xalapa (19°31ʹ55ʺ N; 96°54ʹ35ʺ W) and Coatepec (19°27ʹ08ʺ N; 96°57ʹ41ʺ W) cities in Central Veracruz, México (Fig. 1) during the years 2013–2016, covering all seasons throughout the year (Appendix Table S1). We used point counts on the greenspaces indicated in Fig. 1, and the exact dates of the surveys are provided in Appendix Table S1. Subsequently, we conducted direct feeding observations on all psittacine species detected in six greenspaces of the city of Xalapa during 13 months (June 2017–February 2018, and February 2019–May 2019; Fig. 1). Xalapa is the greenest city of Mexico with vegetation covering 37% of its surface (Falfán et al., 2018).
Figure
1.
Study areas in the cities of Xalapa and Coatepec in Veracruz, México. Census sites are indicated with dots. PEM = Urban Reserve Macuiltepetl, PLS = Urban Park La Señoría, PN = Urban Reserve Natura, CCAD UV = Urban Reserve and Gardens of University of Veracruz, PB = Park Los Berros, JB FJC-SBN = Periurban Reserve Francisco Javier Clavijero-Santuario del Bosque de Niebla, CaCa = Periuran Area in Coatepec Municipality, CaCoa = Urban greenspaces at Los Carriles, CCoa = Coatepec Central Park and Church.
Feeding observations were conducted with binoculars and cameras, during mornings (6:00–11:00) and afternoons (17:00–20:00; Renton, 2001) for a period of 10 days each month, and were duplicated whenever we had less than five records per month. The minimum sampling effort for each greenspace was 80 h per month, for a total of 1040 sampling hours for the whole study. For each sampling day, we did extensive searches in greenspaces and whenever we found a parrot or group of parrots we recorded date, time, location, parrot species, number of birds per species, plant species, and the part of the plant that was consumed. Feeding bouts were recorded every 5 min focusing on the active feeding parrots, including when they changed plant species. When more than one psittacine species was found feeding on the same place, we alternated observations among individuals of the different species.
2.2
Statistical analyses
We used GLMs to determine if the abundance of White-fronted Parrots varied across years and the two surveyed cities. For each analysis we used a negative binomial distribution using the glm.nb function of the MASS library, where the model in R had the form glm.nb (abundance ∼ year + city).
For the diet analyses we only considered those species that had more than 20 feeding bout records. We conducted a standardized pair residual analysis of the observed vs. the expected data to determine if there was a difference in richness of plant species consumed among parrot diets. For this, we used a χ2 test with an α = 0.05, we obtained χ2 values via Jackknife resampling of the relative abundances of each consumed plant species, using a sample size of 22 observations. We also constructed rank-abundance curves (Magurran, 2004) for the plant species consumed by the different parrots and used ANCOVAs to determine if the slopes of the curves were significantly different among the three species of parrots with the highest number of feeding bouts.
For diet breadth we used the Levins standardized index, which quantifies the uniformity of consumed resources by a species in a scale from 0 to 1; 0 indicates a specialized diet and values closer to 1 indicate a generalized diet (Krebs, 1989). Diet overlap among psittacine species was measured using the Pianka index, which also ranges between 0 and 1, where 0 indicates no shared resources and 1 a full overlap in resources between species (Krebs, 1989). In order to determine if feeding overlap is statistically significant, we used a null model keeping constant the observed feeding breadth of each parrot species and conducted 1000 permutations, using the EcoSimR package (Gotelli et al., 2015). Finally, we conducted a non-metric multidimensional scaling (NMDS) analysis to visualize the diet overlap among parrot species across the different sampling months. We used the different months as sampling units and the abundances of the 27 consumed plant species as variables to conduct the ordination. We used the Bray-Curtis index as dissimilarity measure and the analysis was conducted using the “vegan” package in R (Oksanen et al., 2013).
3.
Results
3.1
White-fronted Parrot abundance
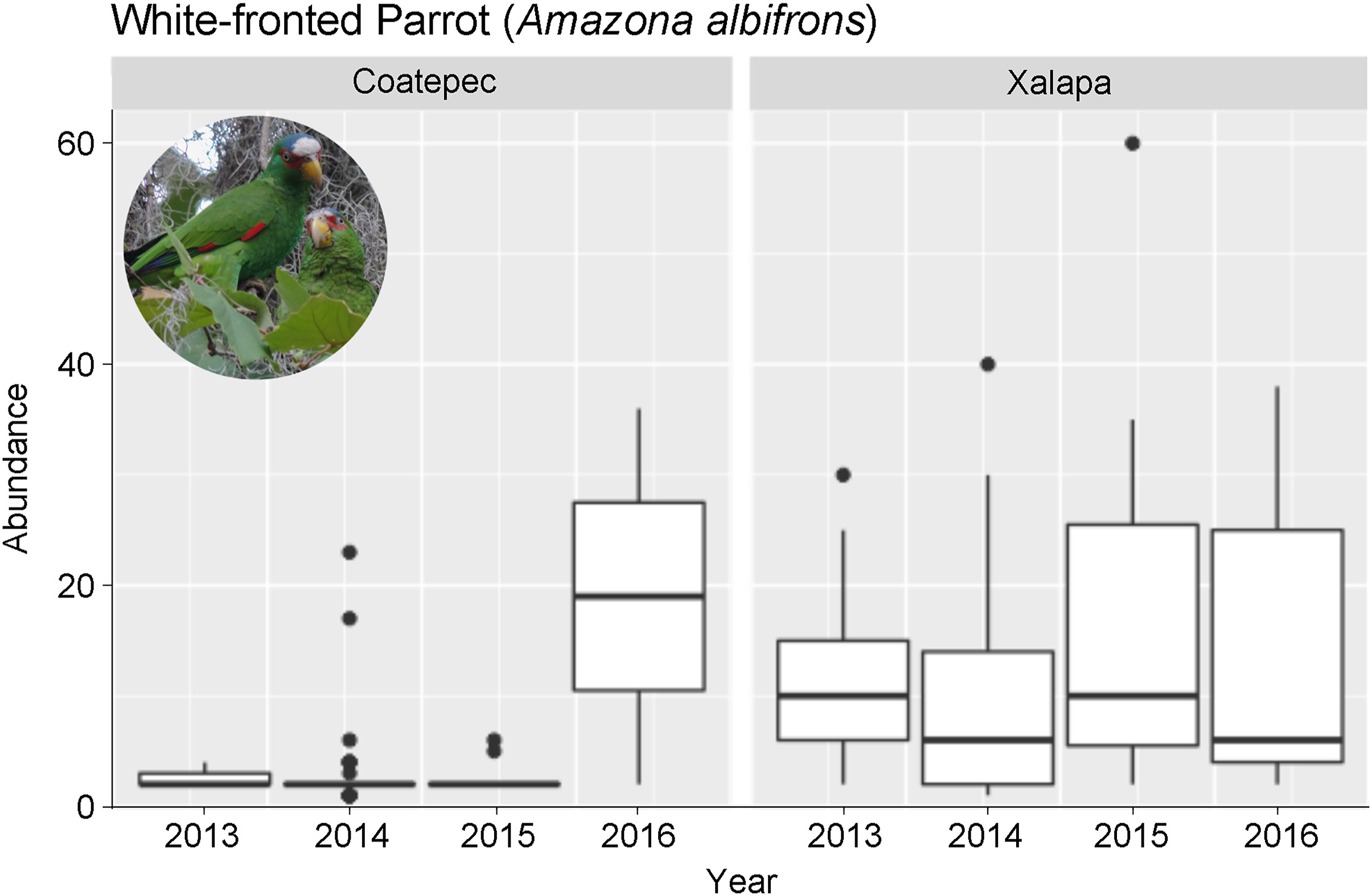
From 2013 to 2016 we conducted a total of 51 censuses in the city of Xalapa and 57 in the city of Coatepec (Appendix Table S1). Xalapa had a significantly higher relative abundance than Coatepec (Table 1). There were no differences in parrot abundances among the four census years in Xalapa, but the year 2016 had significantly higher relative abundances than previous years in Coatepec (Fig. 2).
Table
1.
Results of the GLM analysis for the abundance of the White-fronted Parrot (Amazona albifrons) in the cities of Xalapa and Coatepec.
We obtained a total of 287 feeding bout records, corresponding to 27 plant species and 6 parrot species (Myiopsitta monachus, Eupsittula nana, E. canicularis, Psittacara holochlorus, Pionus senilis, and Amazona albifrons). The White-fronted Parrot had most of the observations (n = 174, 60.6%) and was the species with the highest diet richness (n = 23 plant species), the other parrot species had less than 10 plant species recorded (Appendix Table S2). Non-native plants represented between 30% and 40% of Psittacidae diets (Appendix Table S2). Seeds were one of the most common resources consumed by most parrot species, with the exception of the Green Parakeet (Psittacara holochlorus) that mostly consumed leaves (Appendix Table S3).
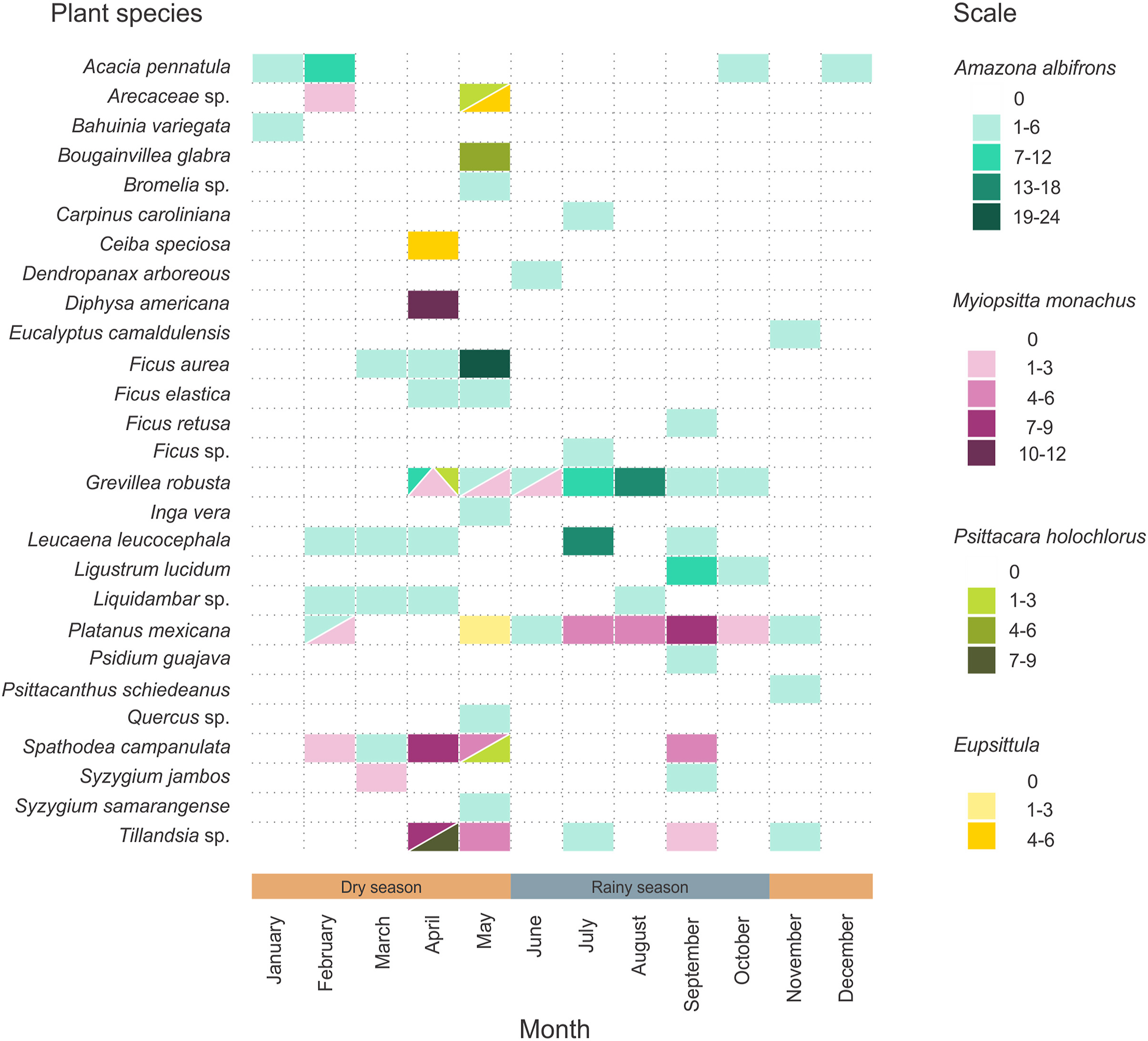
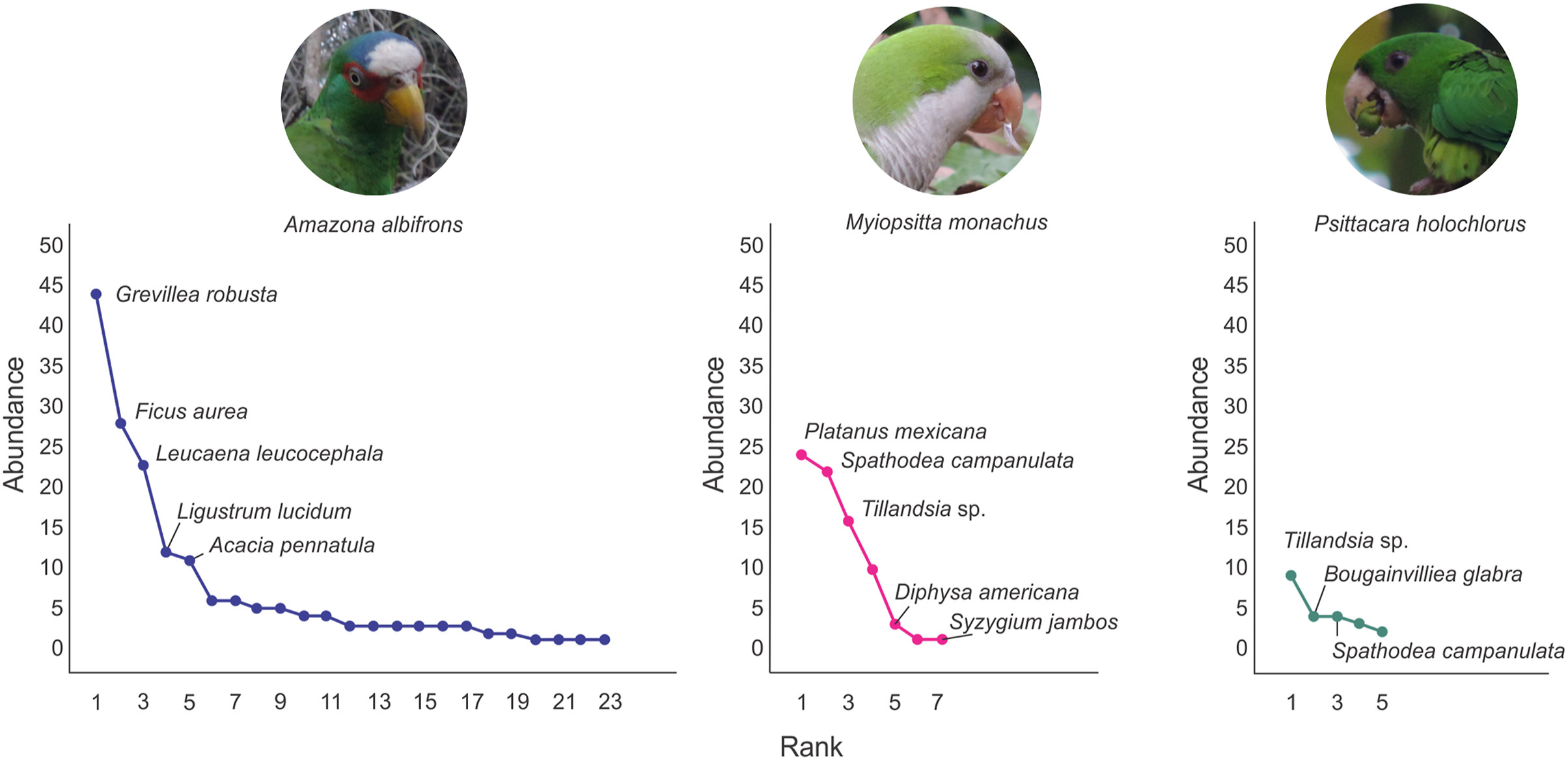
The highest diet richness for all parrots was recorded from November to May. The highest number of feeding bouts was recorded from the end of the dry season (April–May) to the rainy season (June–October; Fig. 3). By the end of the dry season (April–May) we recorded the highest Psittacidae species richness and we also recorded the highest diet overlap among species (Fig. 3). The diets and the slope of the diet abundance curve of the White-fronted Parrots were significantly different to those of the Monk Parakeet (Myiopsitta monachus), but were similar to the diet of the Green Parakeet (Appendix Tables S4 and S5). The diet abundance curve for the White-fronted Parrot showed a steep slope, indicating that few plant species make up most of its diet even when this parrot species had the highest richness in its diet (Fig. 4). The two most important species in the White-fronted Parrot diet were Grevillea robusta (n = 44), followed by Ficus aurea (n = 28). For the Monk Parakeet, the two dominant plant species were Platanus mexicana (n = 24) and Spathodea campanulata (n = 22). The diet of the Green Parakeet was dominated by plants of the genus Tillandsia (n = 9).
Figure
3.
A matrix representation of the different plant species consumed by parrots across different months during June 2017–February 2018, and February 2019–May 2019 in the city of Xalapa, Veracruz, México. The scale (either grey and patterns, or color in online version) indicates the number of times (i.e., feeding bouts) a parrot species consumed the resource. Given that the two parrot species of the genus Eupsittula fed on the same plant species and had the same number of records, we decided to group them as one with the same scale.
The White-fronted Parrot's diet had a lower breadth (Levin's index = 0.32) than the Monk Parakeet (0.52) and the Green Parakeet (0.7), which is explained due to the high dominance by three plant species (54% of records) in the White-fronted Parrot's diet. In terms of diet overlap, the Monk and Green Parakeets shared 56% of their diets, whereas the Monk Parakeet only shared 15% of its diet with the White-fronted Parrot (Appendix Table S6). The null model of diet overlaps among the three species of parrots indicated a non-significant trend to be higher than expected (P = 0.07; Appendix Fig. S1). Regarding diet seasonality, the Monk Parakeet overlaps with the White-Fronted Parrot during the months of February, April, June and September, which is explained by that during those months the Monk Parakeet fed on G. robusta that represents the most important plant in the White-fronted Parrot's diet, and in turn the White-fronted Parrot fed on P. mexicana and Tillandsia sp., both common in the Monk Parakeet's diet (Appendix Fig. S2). Finally, the diet of the Orange-fronted Parakeet (Eupsittula canicularis), the Olive-throated Parakeet (Eupsittula nana), and the White-crowned Parrot (Pionus senilis) do not overlap with those of the White-fronted Parrot and the Monk Parakeet (Appendix Fig. S2), which may only be a result of a low number of observations (Appendix Table S2).
4.
Discussion
In this study we analyzed the relative abundance of the White-fronted Parrot across four years (2013–2016), which is the most common parrot species in the Neotropical cities of Xalapa and Coatepec, México. Furthermore, we analyzed the diet of six parrot species recorded throughout 13 months (2017–2019) in Xalapa. Abundance of the White-fronted Parrot was similar across years, which agrees with results indicating that permanent resident populations in Central Veracruz are the outcome of a recent (i.e., last two decades) natural geographic expansion from southeast México (Mota-Vargas et al., 2020). A previous study detected a diet for the White-fronted Parrot that included 14 plant species (Mota-Vargas et al., 2020), whilst 23 plant species were recorded in this study. The higher plant species richness recorded here may be the result of our more thorough feeding study conducted across 13 months, covering all seasons of a year, which contrasts with the more opportunistic surveys previously conducted. Because the populations that we studied are the result of relatively recent geographical expansions, previous studies might have observed populations in an earlier phase of expansion adapting to novel local resources, contrasting with more permanent and adapted current populations analyzed here. Thus, the White-fronted Parrot is an urban adapter species that has successfully expanded its geographical range both via natural means as well as via anthropogenic introductions in areas such as California, Florida, and Puerto Rico, where it currently has permanent populations (Pranty and Epps, 2002; Mota-Vargas et al., 2020).
The diet of the White-fronted Parrot was dominated by three species: one exotic (G. robusta) and two natives (F. aurea, Leucaena leucocephala), which agrees with what was observed in other studies indicating that parrots consume few plant species, but such resources change in frequency as a function of seasonal availability (South and Pruett-Jones, 2000; Renton, 2001). Some parrots have a specialized diet even during seasons with higher resource availability (Matuzak et al., 2008), like what we found for the White-fronted Parrot that despite having a richer plant species diet, it was the only species with a specialized diet as measured by Levin's index. The low diet overlap of the White-fronted Parrot and the Monk Parakeet indicates that these two species use resources differently in terms of time (seasonality) and space, particularly because the Monk Parakeet was recorded in only one park in Xalapa (see below). Hence, the results from our study confirm that the White-fronted Parrot successfully uses city resources due to its generalist diet and opportunistic behavior, traits shared by many parrot species and successful invasive species (Galetti, 1997; Mack et al., 2000; Muñoz and Real, 2006; Ragusa-Netto and Fecchio, 2006; Boyes and Perrin, 2009; Martens et al., 2013).
The parrot species with the second highest number of feeding records was the non-native invasive Monk Parakeet, which has become a threat for many native bird species, particularly as competition for territories and in some cases lethal encounters with smaller bird species (MacGregor-Fors et al., 2011). This species has communal nests and usually remains close to them given the high maintenance needs of such nests (Bucher et al., 1991), suggesting that their feeding habits would be limited to what is available close to their nests (e.g., maize; Aramburú, 1997). In Xalapa, this parrot species is restricted to one park (La Señoría) feeding mostly on P. mexicana and S. campanulata, that are common around its communal nest. However, when these two species were not producing fruits in the non-breeding season (November–January), the Monk Parakeet was absent probably due to local movements searching for resources in other urban greenspaces (e.g., González-García et al., 2014), but also in periurban and non-urban areas. Although the Monk Parakeet has been previously described as an aggressive species toward native species (Freeland, 1973; Davis, 1974; Long, 1981) we did not record any aggressive behavior toward other parrot or bird species; we even recorded it sharing nesting and feeding sites with other parrot species (e.g., White-fronted Parrot, Green Parakeet). Our observations seem to indicate that the Monk Parakeet has been in the same area within Xalapa city for many years without any further expansion, which contrasts with what has been observed in other locations such as USA and Europe (Clavell et al., 1991; Pruett-Jones et al., 2012). It is possible that an ecological factor limits this parrot species’ invasive potential in Xalapa; or, it may be in a pre-expansion phase in the process of invasion (Aagaard and Lockwood, 2014). Furthermore, the Monk Parakeet was nesting on an introduced plant, agreeing with previous studies (Muñoz-Jiménez and Alcántara-Carbajal, 2017). Thus, we suggest that city authorities should avoid the use of ornamental exotic species that favor the establishment of the Monk Parakeet. Specifically, for Xalapa we refer to plant species such as Araucaria araucana, Phoenix canariensis, Washingtonia robusta, and Syagrus romanzoffiana (González-García et al., 2016).
We recorded the Green Parakeet only during the last months of the dry season (April–May 2019) probably because its diet is low in plant species richness. During this period of the year resources are scarcer in more natural environments due to reduced precipitation (e.g., Renton, 2001; Davis et al., 2012, 2015), which is why this species was recorded within the city feeding on species such as Tillandsia sp., Bougainvilliea glabra and S. campanulata. Because of the low number of observations of Green Parakeets in only one greenspace, we are not able to rule out that the presence of this parrot species was due to an escape from captivity. Moreover, this species is poorly known in urban areas and the few studies in cities from the USA have recorded the Green Parakeet feeding on wasp larvae and Lagerstroemia indica seeds (Alexander, 2016; Graves, 2018). In the case of the other three parrot species, we also recorded a low number of observations that may be due to their rarity within the city and across the year, which would classify them as urban utilizers. Therefore, it would be important to study these parrot species in the peri-urban area in order to find out if they are avoiding the city's greenspaces, and if they are to identify the factors that determine it (e.g., competition with the White-fronted Parrot?).
Importantly, exotic plant species represented ∼41% of recorded feeding bouts for all parrot species in this study. This may simply reflect the fact that native vegetation in Xalapa represents only 36%, whereas introduced plant species are ∼55% of the woody vegetation (Falfán and MacGregor-Fors, 2016). Additionally, introduced plant species have longer flowering and fruiting periods, and in some cases a higher caloric content (e.g., S. campanulata; Bahadur et al., 1986; Rangaiah et al., 2004; Silva et al., 2015). In this sense, success of both native and introduced parrot species in urban environments is largely due to their behavioral and feeding flexibility (Pruett-Jones et al., 2012; Mori et al., 2017; Ramírez-Albores and Aramburú, 2017), which allows them to successfully exploit exotic plant species during seasons when resources are scarce in their natural environments (e.g., Renton, 2001; Smith and Lill, 2008; Silva, 2018).
Why was there the highest overlap among parrot species by the end of the dry season? How did exotic plant species (particularly in Xalapa City) impact parrots diet and coexistence of three species? A possible explanation for the overlap in the diet of the different species of parrots could be related to the phenology of the plants they consume, since it can be observed that at the end of the dry season, which corresponds to spring, and the beginning of the summer, many of the plant species have flowers, fruits or both, which are resources that are part of the diet of the different species of parrots studied; for example, Grevillea robusta flowers from May to July and produces fruits from July to August, or Spathodea campanulate that flowers from January to July (Appendix Table S7). Alternatively, there are species of exotic plants (e.g. Eucalyptus camaldulensis) that due to their high plasticity, can proliferate and develop flowers or fruits practically throughout the year, which implies food resources for birds in periods in which native plants (e.g. Carpinus carolineana, Platanus mexicana) do not provide such resources. Another important factor is that the fruits of some exotic plants are part of the diet of some birds, as was observed in this work; also, previous studies have shown that even the fruits of exotic species are more attractive compared to the fruits of native species, which may be due to availability, quantity, or being more attractive due to their size (LaFleur et al., 2007; Jordaan and Downs, 2012; Gray and Heezik, 2016). Some exotic plants such as Syzygium samarangense or S. jambos described in this work, have large and fleshy fruits, which are attractive to birds and represent an important source of food for parrots. Summarizing, our results agreed with our initial expectations that the White-fronted Parrot – the largest parrot in the study – has a larger diet breadth than the other parrot species, and that it is present year-round with similar abundances across years. And contrary to expectations, the diet overlap between the White-fronted Parrot and the similar sized Monk Parakeet was low, which is likely due to the restricted spatial distribution of the exotic Monk Parakeet and to the different preferences of plant species as reflected in their rank-abundance curves.
Author contributions
CÁ-C conceived the idea of the psittacine diet and CM-V the idea of the abundance of psittacines. CM-V, DS-A, IM-F and SLAW helped design the study. CÁ-C and CM-V conducted field work and collected data. CÁ-C, CM-V and DS-A analyzed and interpreted the data. CÁ-C and DS-A wrote and revised the paper. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank A. Sandoval-Comte for preparing the maps in Fig. 1, some figures were improved using Biorender.com. Thanks also go to K. P. Parra-Noguez, P. Téllez-Mendoza and Stéphanie Baltazar-Hernández for their support in the A. albifrons field work. The research was supported by the Consejo Nacional de Ciencia y Tecnología (CONACYT) who provided study grant (450578) to CÁ-C to conduct her master's studies at INECOL, A.C. DS-A was supported by CONACYT program Problemas Nacionales project number 2015-01-1628.
Aagaard, K., Lockwood, J., 2014. Exotic birds show lags in population growth. Divers. Distrib. 20, 547-554.
Adler, F.R., Tanner, C.J., 2013. Urban Ecosystems: Ecological Principles for the Built Environment, first ed. Cambridge University Press, Cambridge.
Alexander, C., 2016. Green Parakeets feeding on Wool-bearing Gall-wasp larvae (Andricus quercuslanigera). Bull. Texas Ornithol. Soc. 49, 100-102
Aramburu, R.M., 1997. Ecologia alimentaria de la cotorra (Myiopsitta monachus) en la provincia de Buenos Aires, Argentina (Aves: Psittacidae). Physis. Seccion C. 53, 29-32
Aronson, M.F.J., La Sorte, F.A., Nilon, C.H., Katti, M., Goddard, M.A., Lepczyk, C.A., et al., 2014. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. Biol. Sci. 281, 20133330.
Bahadur, B., Chaturvedi, A., Swamy, N.R., 1986. Nectar types in Indian plants. Proc. Indian Acad. Sci. 96, 41-48.
Berkunsky, I., Quillfeldt, P., Brightsmith, D.J., Abbud, M.C., Aguilar, J.M.R.E., Aleman-Zelaya, U., et al., 2017. Current threats faced by Neotropical parrot populations. Biol. Conserv. 214, 278-287.
Bonier, F., Martin, P.R., Wingfield, J.C., 2007. Urban birds have broader environmental tolerance. Biol. Lett. 3, 670-673.
Boyes, R.S., Perrin, M.R., 2009. The feeding ecology of Meyer’s Parrot Poicephalus meyeri in the Okavango Delta, Botswana. Ostrich 80, 153-164.
Bucher, E.H., Martin, L.F., Martella, M.B., Navarro, J.L., 1991. Social behavior and population dynamics of the monk parakeet. Proc. Int. Ornithol. Congr. 20, 681-689
Clavell, J., Martorell, E., Santos, D.M., Sol, D., 1991. Distribucio de la Cotorreta de Pit Gris Myopsitta monachus a Catalunya. Butll. G. C. A. 8, 15-18.
Davis, A., Major, R.E., Taylor, C., 2012. Seasonal abundance and habitat use of Australian parrots in an urbanized landscape. Landsc. Urban Plan. 106, 191-198.
Davis, A., Major, R.E., Taylor, C., 2015. The association between nectar availability and nectarivore density in urban and natural environments. Urban Ecosyst. 18, 503-515.
Davis, L.R., 1974. The Monk Parakeet: a potential threat to agriculture. In: Johnson, W.V., Marsh, R.E. (Eds.), Proceedings of the 6th Vertebrate Pest Conference. University of California, Riverside, pp. 253–256.
Falfan, I., MacGregor-Fors, I., 2016. Woody neotropical streetscapes: a case study of tree and shrub species richness and composition. Madera y Bosques 22, 95-110
Falfan, I., Munoz-Robles, C.A., Bonilla-Moheno, M., MacGregor-Fors, I., 2018. Can you really see ‘green’? Assessing physical and self-reported measurements of urban greenery. Urban For. Urban Green. 36, 13-21.
Fischer, J.D., Schneider, S.C., Ahlers, A.A., Miller, J.R., 2015. Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv. Biol. 29, 1246-1248.
Freeland, D.B., 1973. Some food preferences and aggressive behavior by Monk Parakeets. Wilson Bull. 85, 332-334
Galetti, M., 1997. Seasonal abundance and feeding ecology of parrots and parakeets in a lowland Atlantic forest of Brazil. Ararajuba 5, 115-126
Gascon, C., Lovejoy, T.E., Bierregaard, Jr. R.O., Malcolm, J.R., Stouffer, P.C., Vasconcelos, H.L., et al., 1999. Matrix habitat and species richness in tropical forest remnants. Biol. Conserv. 91, 223-229
Gonzalez-Garcia, F., Straub, R., Lobato, G.J.A., MacGregor-Fors, I., 2014. Birds of a neotropical green city: an up-to-date review of the avifauna of the city of Xalapa with additional unpublished records. Urban Ecosyst. 17, 991-1012
Gonzalez-Garcia, F., Straub, R., Lobato, G.J.A., MacGregor-Fors, I., Santiago Alarcon, D., 2016. Nuevos registros y notas adicionales comentadas sobre la avifauna de la ciudad de Xalapa, Veracruz, Mexico. Acta Zool. Mex. 32, 253-269
Gotelli, N.J., Hart, E.M., Ellison, A.M., 2015. EcoSimR: null model analysis for ecological data. R package version 0.1.0. .
Graves, G.R., 2018. Avian feeding on seed of the exotic ornamental Lagerstroemia indica (Crapemyrtle). Southeast. Nat. 17, 293-298.
Gray, E., van Heezik, Y., 2016. Exotic trees can sustain native birds in urban woodlands. Urban Ecosyst. 19, 315-329
Jordaan, L.A., Downs, C.T., 2012. Nutritional and morphological traits of invasive and exotic fleshy-fruits in South Africa. Biotropica 44, 738-743
Kark, S., Iwaniuk, A., Schalimtzek, A., Banker, E., 2007. Living in the city: can anyone become an ‘urban exploiter'? J. Biogeogr. 34, 638-651
Krebs, C.J., 1989. Ecological Methodology. Harper & Row, New York.
LaFleur, N.E., Rubega, M.A., Elphick, C.S., 2007. Invasive fruits, novel food and choice: an investigation of European Starling and American Robin frugivory. Wilson J. Ornithol. 119, 429-438
Long, J.L., 1981. Introduced Birds of the World. Universe Books, New York.
MacGregor-Fors, I., Calderon-Parra, R., Melendez-Herrada, A., Schondube, S., 2011. Pretty, but dangerous! Records of non-native Monk Parakeets (Myiopsitta monachus) in Mexico. Rev. Mex. Biodiv. 82, 1053-1056
Mack, R.N., Simbeloff, D., Londsdale, W.M., Evans, H., Clout, M., Bazzaz, F.A., 2000. Biotic invasions: causes, epidemiology, global consequences, and control. Ecol. Appl. 10, 689-710
Martens, J., Hoppe, D., Woog, F., 2013. Diet and feeding behaviour of naturalized Amazon Parrots in a European city. Ardea 101, 71-76
Matuzak, G.D., Bezy, B., Brightsmith, D.J., 2008. Foraging ecology of parrots in a modified landscape: seasonal trends and introduced species. Wilson J. Ornithol. 20, 353-365
Maxwell, S.L., Fuller, R.A., Brooks, T.M., Watson, J.E., 2016. Biodiversity: the ravages of guns, nets and bulldozers. Nature 536, 143-145
Menchetti, M., Mori, E., 2014. Worldwide impact of alien parrots (Aves Psittaciformes) on native biodiversity and environment: a review. Ethol. Ecol. Evol. 26, 172-194
Mori, E., Grandi, G., Menchetti, M., Tella, J.L., Jackson, H.A., Reino, L., et al., 2017. Worldwide distribution of non-native Amazon parrots and temporal trends of their global trade. Anim. Biodivers. Conserv. 40, 49-62
Mota-Vargas, C., Parra-Noguez, K.P., Rojas-Soto, O., 2020. Analisis del conocimiento historico de la distribucion geografica y ecologica del loro frente blanca, Amazona albifrons, con evidencia de colonizacion reciente. Rev. Mex. Biodivers. 91, 1-13
Munoz, A.R., Real, R., 2006. Assessing the potential range expansion of the exotic monk parakeet in Spain. Divers. Distrib. 12, 656-665
Munoz-Jimenez, J.L., Alcantara-Carbajal, J.L., 2017. La cotorra argentina (Myiopsitta monachus) en el Colegio de Postgraduados: ¿una especie invasiva? Huitzil 18, 38-52
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., O’hara, R.B., et al., 2013. Package ‘vegan’. Community Ecology Package, Version. 2, pp. 1–295.
Pranty, B., Epps, S., 2002. Distribution, population, status, and documentation of exotic parrots in Broward County Florida. Florida Field Nat. 30, 111-131
Pruett-Jones, S., Appelt, C.W., Sarfaty, A., van Vossen, B., Leibold, M.A., Minor, E.S., 2012. Urban parakeets in Northern Illinois: a 40-year perspective. Urban Ecosyst. 15, 709-719
Ragusa-Netto, J., Fecchio, A., 2006. Plant food resources and the diet of a parrot community in a gallery forest of the southern Pantanal (Brazil). Braz. J. Biol. 66, 1021-1032
Ramirez-Albores, J.E., Aramburu, R.M., 2017. De Suramerica para Mexico, la invasion de la cotorra argentina (Myopsitta monachus Baddaert, 1783). Rev. Biodivers. Neotrop. 7, 86-97
Rangaiah, K., Purnachandra Rao, S., Solomon Raju, A.J., 2004. Bird-pollination and fruiting phenology in Spathodea campanulata Beauv. (Bignoniaceae). Beitr. Biol. Pflanzen. 73, 395-408
Renton, K., 2001. Lilac-crowned Parrot diet and food resource availability: resource tracking by a parrot seed predator. Condor. 103, 62-69
Shanahan, D.F., Strohbach, M.W., Warren, P.S., Fuller, R.A., 2014. The challenges of urban living. In: Gil, D., Brumm, H. (Eds.), Avian Urban Ecology Behavioural and Physiological Adaptations. Oxford University Press, Oxford, pp. 3–20.
Silva, P.A., 2018. Massive consumption of unripe slash pine (Pinus elliottii) seeds by blue-and-yellow macaws (Ara ararauna). Ornitol. Neotrop. 29, 301-308
Smith, J., Lill, A., 2008. Importance of eucalypts in exploitation of urban parks by Rainbow and Musk Lorikeets. Emu 108, 187-195
Sol, D., Bartomeus, I., Gonzalez-Lagos, C., Pavoine, S., 2017. Urbanisation and the loss of phylogenetic diversity in birds. Ecol. Lett. 20, 721-729
South, J.M., Pruett-Jones, S., 2000. Patterns of flock size, diet, and vigilance of naturalized Monk Parakeets in Hyde Park, Chicago. Condor 102, 848-854

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds