Johann H. VAN NIEKERK. 2013: Vocal structure, behavior and partitioning of all 23 Pternistis spp. into homologous sound (and monophyletic) groups. Avian Research, 4(3): 210-231. DOI: 10.5122/cbirds.2013.0020
Citation:
Johann H. VAN NIEKERK. 2013: Vocal structure, behavior and partitioning of all 23 Pternistis spp. into homologous sound (and monophyletic) groups. Avian Research, 4(3): 210-231. DOI: 10.5122/cbirds.2013.0020
Johann H. VAN NIEKERK. 2013: Vocal structure, behavior and partitioning of all 23 Pternistis spp. into homologous sound (and monophyletic) groups. Avian Research, 4(3): 210-231. DOI: 10.5122/cbirds.2013.0020
Citation:
Johann H. VAN NIEKERK. 2013: Vocal structure, behavior and partitioning of all 23 Pternistis spp. into homologous sound (and monophyletic) groups. Avian Research, 4(3): 210-231. DOI: 10.5122/cbirds.2013.0020
Department of Environmental Sciences, College of Agriculture and Environmental Sciences, University of South Africa, PO Box 392, Pretoria, 0003, South Africa
The aims of this research were (1) to provide a description of spurfowl Pternistis spp. calls and their social context; (2) to describe the divergence of advertisement calls; and (3) to appropriate 23 spurfowl species to homologous sound groups which have been synthesized with recognized monophyletic groups within Pternistis spurfowls. Sound group partitioning was primarily based on male advertisement calls. A total of 218 recordings (rendering ~300 identifiable calls) were analyzed covering 22 out of 23 spurfowl species in Africa. One species was assessed from written accounts. The repertoire size per spurfowl varies between 7 and 11 calls. Spurfowl calls were arranged into three broad categories including (1) advertisement calls; (2) maintenance calls including distress calls, juvenile whining ("mews"), cheeps and comfort calls; and (3) male-female and female-offspring bonding calls. Spurfowl species were set out in eight sound groups of which five were more or less congruent with the monophyletic groups of Hall (1963), but sound groups produced more partitioning as Hall described only five groups relevant to Pternistis spp. The divergence of advertisement calls apparently minimizes hybridization between sympatric species but the "genetic distance" between spurfowl species is relatively small causing hybridization among spurfowl species. Despite the vocalizations of Hartlaub's Spurfowl (P. hartlaubi) differing significantly from the rest of the spurfowls, sound analyses suggest that it remains within Pternistis.
Hall (1963) regarded all spurfowls and francolins as one genus (Francolinus) (Order: Galliformes, Family: Phasianidae). However, the phylogeny of the "francolins"
(sensu Francolinus) has been overhauled by Crowe et al.
(1992) and supported by Bloomer and Crowe (1998), who based their evidence on morpho-behavioral and DNA analysis, dividing it into two clades, the francolins (Francolinus, Dendroperdix, Scleroptila, Peliperdix) and spurfowls (Pternistis) (also see Little and Crowe, 2011). This split was also supported by syringeal studies that demonstrate differences between the two clades (Mandiwana-Neudani et al., 2011). Pternistis spurfowls fall within five of Hall's (1963) putative monophyletic groups that include the vermiculated, bare-throated, scaly and Montane groups. Furthermore, the vermiculated group was divided into a northern and southern vermiculated group.
Few published data exist on the vocal behavior of Pternistis. Apart from Hartlaub's Spurfowl (P. hartlaubi)
(Komen, 1987), one or two calls have been described for each species of Pternistis in standard reference books (e.g. Madge and McGowan, 2002; Hockey et al., 2005; Little and Crowe, 2011). Milstein and Wolff (1987) compared one or two calls of spurfowls (Pternistis) with francolin calls (Dendroperdix, Peliperdix and Scleroptilia) and came to a valid conclusion that francolins make tonal whistles while spurfowls produce atonal raucous calls.
Generally, sounds are important for distinguishing spurfowl species in the wild and also for separating spurfowls taxonomically from closely related species, such as the francolins, because the clades require different conservation measures (Milstein and Wolff, 1987). In South Africa it is not uncommon to find five spurfowl and francolin species living together on the same farm in close proximity (300–2000 ha) (J.H. van Niekerk, unpublished). Furthermore, spurfowls are actively hunted in Africa (Viljoen, 2005), and calls can therefore also be used to entice spurfowl responses by using sound-playbacks for management purposes, such as locating territories, sexing, and determining densities and habitat preferences (cf. Wolff, 1978; Little and Crowe, 1992; Fuller et al., 2012; van Niekerk, 2012).
The aims of this research were (1) to provide a detailed description of spurfowl calls and their social context; (2) to describe the divergence of advertisement calls; and (3) to appropriate spurfowl species to homologous sound groups based primarily on male advertisement calls and correlated with the recognized monophyletic groups.
Methods
Of the 23 spurfowl species in Africa, six occur in southern Africa (Hockey et al., 2005; Little and Crowe, 2011), and these were observed with the aid of binoculars in the field from 2002 to 2012 to understand each call in a social context (Table 1). Nahan's Partridge (Ptilopachusnahani), previously regarded as a spurfowl (Sinclair and Ryan, 2003; Fuller et al., 2012), is no longer a spurfowl and is therefore excluded from Table 1 (Crowe, 2010). The sound recordings made by the author were undertaken during research for published papers, but most calls have not yet been published (van Niekerk et al., 2009; van Niekerk, 2004, 2010, 2011, 2012). Of particular importance is that the social context of calls (group size, courtship behavior, care of offspring and stressful situations), and the responses of conspecifics, were noted during natural calls/sound-playbacks.
Table
1.
List with Pternistis spurfowls and calls assessed, source of information and time spent observing southern African spurfowls species during fieldwork
Species
Number of recordings assessed and time spent observing southern African spurfowls
Source (and locality coordinates in the case of southern African spurfowls)
All the species occurring in southern Africa listed in Table 1 (south of the Cunene, Okavango and Zambezi Rivers) were enticed with sound-playbacks that had been pre-recorded at the locality where the birds were observed. Similar calls from different individuals but on different tracks were used during playbacks. The target spurfowls were approached on foot and from a vehicle. When the latter was used the vehicle was brought to a standstill by switching it off. Males were differentiated by noting the spurs on their legs. A period of 5 minutes (encompassing playback time) was allowed for spurfowls to respond during or just after every playback. As the spurfowls responded they were watched with a pair of binoculars to note general behavior and to differentiate between different spurfowls calling. The distances between the target bird(s) and the recorder or soundplayback device and the bird(s) were 1–5 m for feeding and male-female bonding calls, 20–30 m for alarm calls and 25–35 m for advertisement and other loud calls, such as distress calls. Two Sony recorders (ICD-BX800) were used, one for sound-playback and another for recording purposes. The recorders were held/placed about 1 m above ground level and were often left in that position for one hour of recording but during traversing the instruments embedded microphone was pointed in the direction of spurfowls.
A relative assessment of volume was also described. This was based mainly on the southern African field studies but a relative assessment could also be assessed where different calls were recorded simultaneously on one soundtrack for species occurring in the rest of Africa. Volumes of the southern African species was identified as either low (~50–65 dB), medium (~65–95 dB) or high (~95–120 dB), mainly based on general sound level recordings made with the aid of a Lutron SL-4013 meter at 25–35 m from the bird rather than direct volume (dB) readings from Raven Pro 1.4. However, long exposure to many species in the field provided the author with a good basis to arrange the different calls into relative volume categories.
Table 1 also shows the calls that were obtained from other sources, comprising those spurfowl calls that were available for species occurring in the rest of Africa. This material was augmented with call and behavioral descriptions from the existing literature (e.g. Madge and McGowan, 2002; Hockey et al., 2005). All the calls used for the outgroup analysis were obtained from www.xeno-canto.org.
Spectrogram analysis
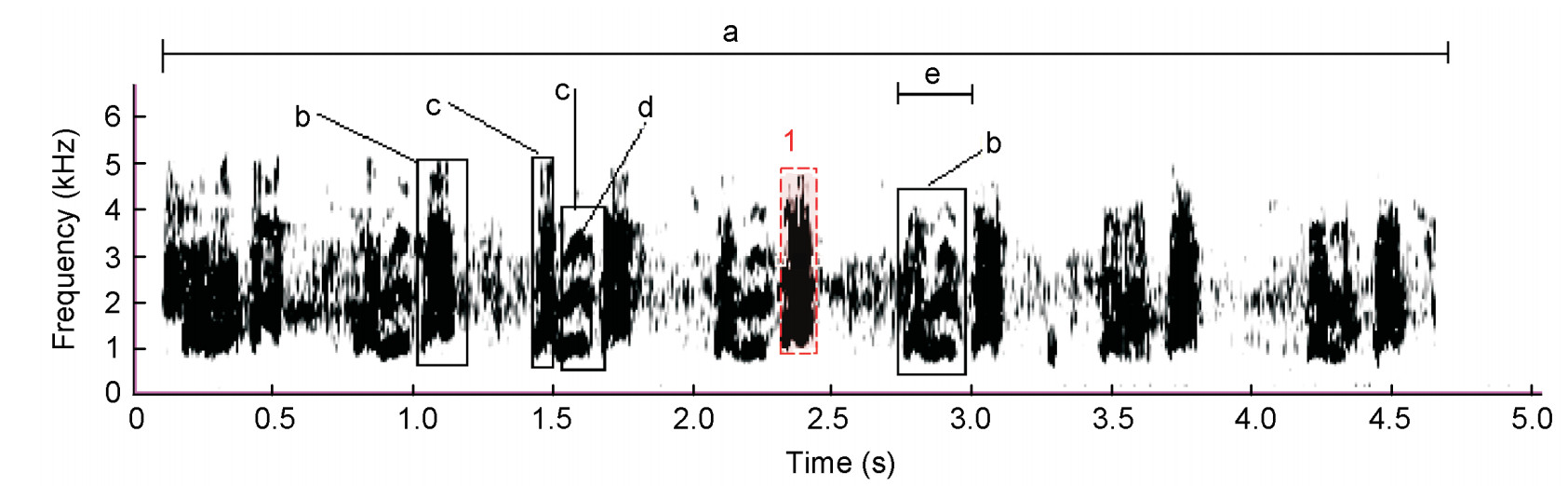
Calls were imported to Raven Pro 1.4 to visualize spectrograms for comparative analysis. Spectrogram parameters were fixed at sample size = 256, overlap = 50%, time grid spacing = 128 and discrete Fourier transformation = 256. A Hanning window type was used (Charif et al., 2010). The sampling rate for the southern African birds was 44.10 kHz. The syllable (Fig. 1) was used as the basic unit for sound/statistical comparison between call types.
Figure
1.
Measurement of spectrograms: a = duration of call, b = syllable, c = element of a syllable, d = harmonic, e = duration of syllable, 1 = selection box around a syllable to determine frequency bandwidth and duration
Selection of spectrogram measurements to describe calls
Six standard spectrogram measurements were selected (Raven Pro 1.4, Charif et al., 2010) which were deemed important to differentiate between high- and low-intensity guttural crows and between alarm calls on the ground and during flight, including (1) duration of call (seconds), (2) delta time of syllable (length of syllable measured in seconds), (3) high frequencies (kHz), (4) low frequencies (kHz), (5) delta frequencies (bandwidth in kHz), and (6) centre frequencies (kHz). The selection boxes which were drawn around syllables (function in Raven Pro 1.4) to determine spectrogram measurements (Fig. 1) were done simultaneously with audible sound in order to standardize the demarcation of frequency bandwidth (vertical measurements in kHz) and duration (horizontal measurements in seconds), that is to limit the blank areas (no tracings) within the selection boxes (cf. Erbe and King, 2008 and A. Warde inlitt.). In addition, means and standard variation statistics were provided (Table 2).
Table
2.
Comparative statistics of crows and alarm calls found in Pternistis spurfowls, deployed against six spectrogram measurements
High intensity guttural crow
Low intensity crow
Alarm call on ground
Alarm call in flight
Total number of calls analyzed (species)
18
13
14
8
(P. swainsonii, P.afer, P. adspersus, P.rufopictus, P. leucoscepus)
(P. swainsonii, P.afer, P. capensis, P.natalensis, P. erckelii, P. bicalcaratus, P. leucoscepus, P.squamatus)
(P. swainsonii, P.afer, P. adspersus, P.capensis, P. natalensis, P.erckelii, P. leucoscepus, P. hildebrandti, P.ochropectus)
(P. swainsonii, P.afer, P. adspersus, P. capensis, P.natalensis)
An element is part of a syllable, for example "kowaak" is one syllable but part of the syllable starts with a low frequency but ends with a higher frequency (upsweep). The low and high frequencies form two elements of the same syllable (Fig. 1).
Differentiation between crows and non-crows
A loud advertisement crow was designated to one of two crow types that produced either (1) guttural syllables which was sometimes mixed with tonal syllables and (2) squelching syllables, which were often mixed with laughing syllables (Table 3). Where more than one of these crow types occurred in one call, the dominant one (> 50% of the call duration) was used for designation to one of the crow types.
Table
3.
Calls of Pternistis spurfowls based on homologous sound principles (spectrogram structure) arranged in sound groups which were also compared with existing phylogenetic results for congruence/incongruence. Numbers between parentheses indicate sample sizes.
"??" = tentatively placed and "?" = further field work required.
Conversely, a non-crow call was mainly tonal (Table 3), and generally possessed a horizontal solid spectrogram texture (narrower frequency bandwidth that possessed fewer harmonics) as against the vertically oriented blurry-perforated texture of all the crow calls.
Sound groups
A sound group is two or more species that share sound characteristics (e.g. homology of spectrogram structure as in Fig. 1), or a single species with unique characteristics that set it apart from the rest of the spurfowls based on a coherent systematic approach (cf. Newton, 2003). Sound groups were mainly restricted to loud advertisement calls that address one of the aims of this paper that these calls diverged more than other call categories (cf. Deregnaucourt, 2010). Where species also possessed duets as advertisement calls (n = 4), the non-duet advertisement calls were used as the primary call for sound group partitioning since all other species (~19) possessed the latter and not a duet (Table 3). However, partitioning below the crow types (i.e. guttural, squelch, and guttural with tonal elements) was also conducted (see sound groups) based on non-crow advertisement calls (Table 3). Sister species refer to species that share an immediate ancestor not shared with another closely related species (cf. Newton, 2003).
Hall's groups were used as a "platform" from which congruence or incongruence between her groups and sound groups was determined (Table 3). These groups were also used by Madge and McGowan (2002) and are regarded as the most comprehensive (partitioning all species) treatment available. Having done this, the sound groups were synthesized with morphological and bio-geographical criteria and a limited DNA component added by Bloomer and Crowe (1998). A selection of outgroup species was chosen from the studies of Kimball et al. (2011) to identify primitive sound character states and polarity.
Results
A total of 218 recordings (rendering ~300 identifiable calls) were analyzed covering 22 of the 23 spurfowl species in Africa (Table 1). No direct recordings could be found for Jackson's Spurfowl (P. jacksoni) but a description of its call was given by Madge and McGowan (2002). Spurfowl calls were classified into at least three broad categories: (1) advertisement, (2) maintenance and (3) bonding calls.
Category 1: Advertisement calls
This category consists of advertisement crows (Fig. 2) and advertisement calls without crows (Table 3). In all the southern African species as well as the Djibouti Spurfowl (P. ochropectus), these advertisement calls are territorial calls (M. Crainger, in litt.). In other words, males demarcate territories from which other males are kept out but also attract females (cf. van Niekerk, 2009, 2011; J.H. van Niekerk, unpublished).
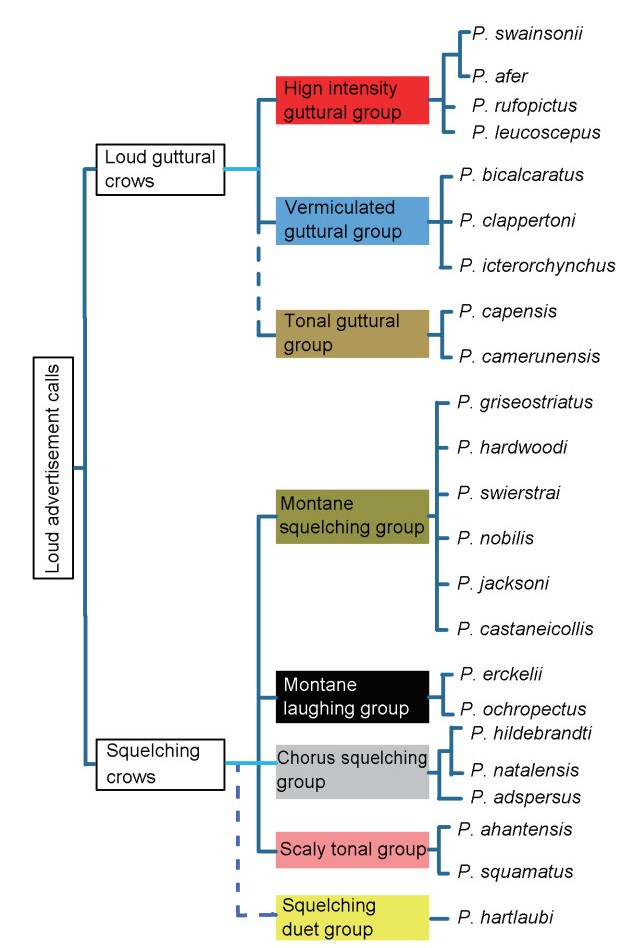
Figure
2.
Divergence of loud advertisement crows of Pternistsis spurfowls deployed in eight recognizable sound groups. Colored blocks correspond with sound groups in Table 3. Dotted line = unresolved.
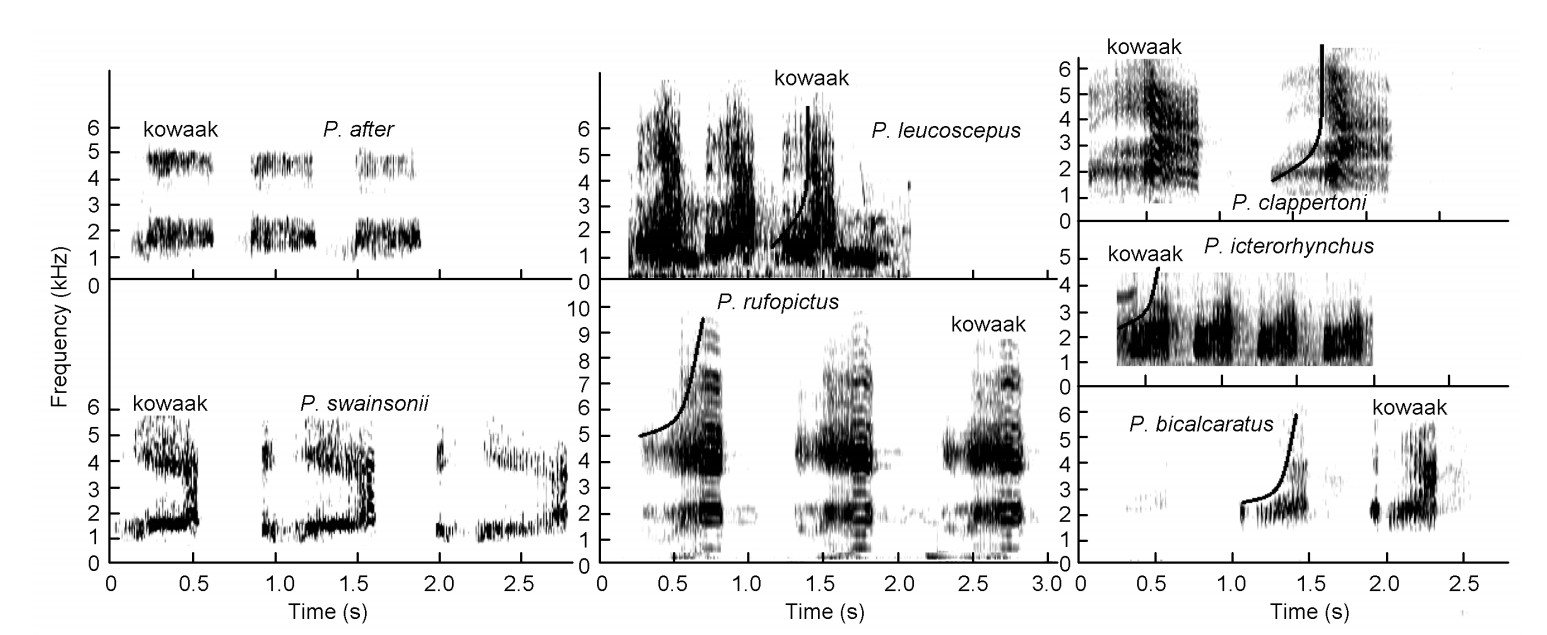
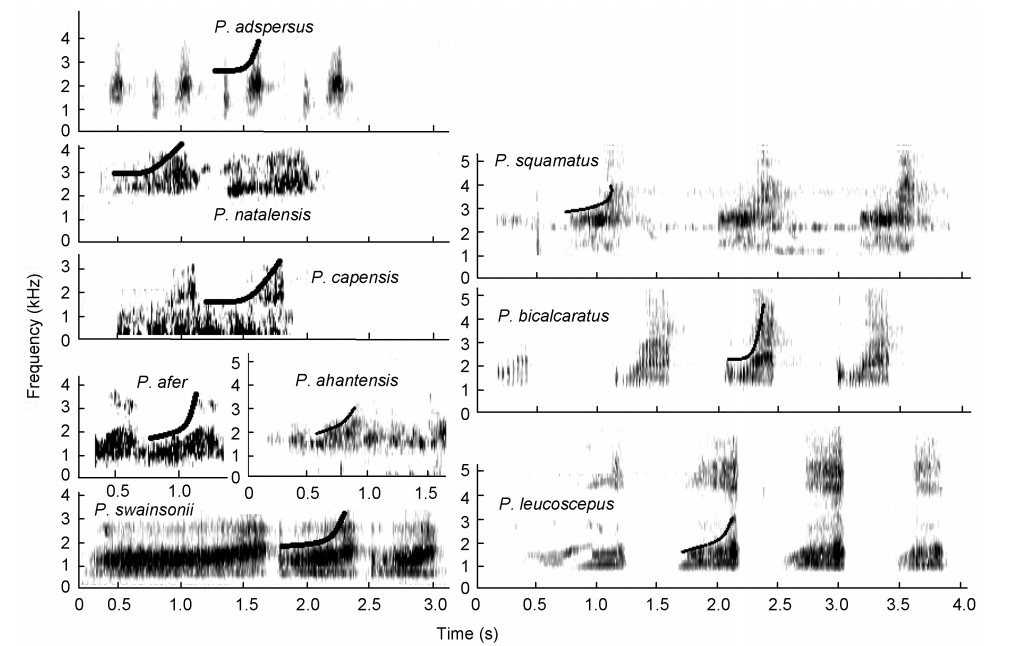
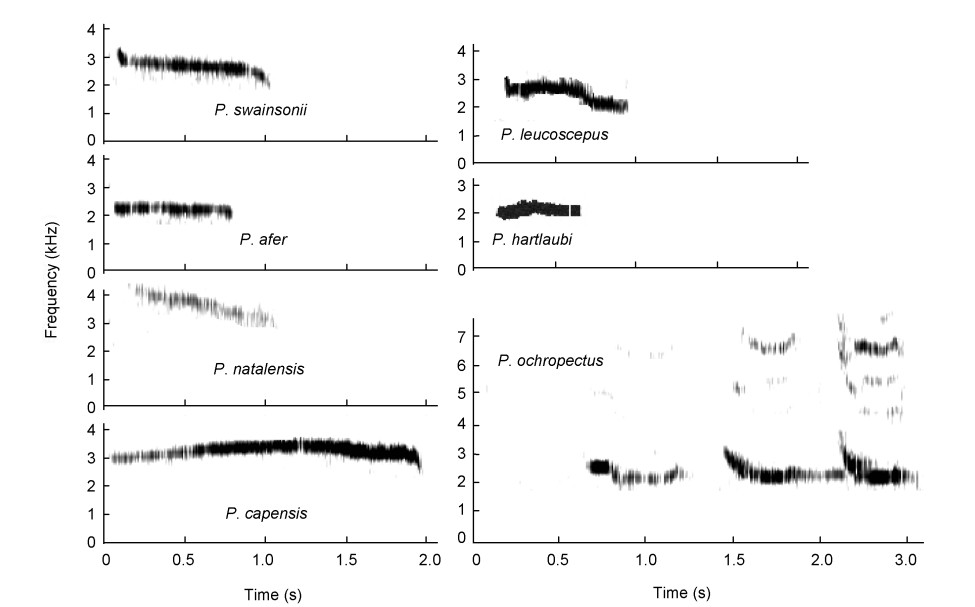
This loud call is a drawn-out, guttural, atonal crow ("kowaak") with three to four similar syllables in terms of volume, length of syllable and pitch, in succession in one call (Figs. 2 and 3, and Table 3). High-intensity guttural crows consist of two sound groups:
Figure
3.
High-intensity guttural crows of Pternistis spurfowls visualized on spectrograms. The black curve on the graph shows an upsweep tendency of the syllable (from low to higher frequency). Note the distinctive vertically oriented blurry-perforated texture.
(1) the bare-throated guttural group (Fig. 2) that invariably turn their heads vertically upwards to inflate their bare throats during the loud advertisement crow to maximize their presence visually and vocally. Skypointing was not observed during the low-intensity guttural crows (see later). Young bare-throated spurfowl males of Red-necked (P. afer) and Swainson's Spurfowls only start crowing when the red coloration appears on their throats. Swainson's and Red-necked Spurfowls are sister species and could be retained as such (Bloomer and Crowe, 1998) and;
(2) the vermiculated guttural group that possess fully-feathered throats with an upsweep syllable in all cases (Table 3, Figs. 2 and 3). The sky-pointing behavior of the bare-throated spurfowls was not recorded in the fully-feathered throated spurfowls (Madge and McGowan, 2002). These two sound groups have similar sound structures but based on the studies of Hall (1963) and Madge and McGowan (2002), which placed these spurfowls into two different groups (Table 3), partitioning of the two sound groups of Hall was not in conflict with the groups and was retained as such. These two groups form sister sound groups.
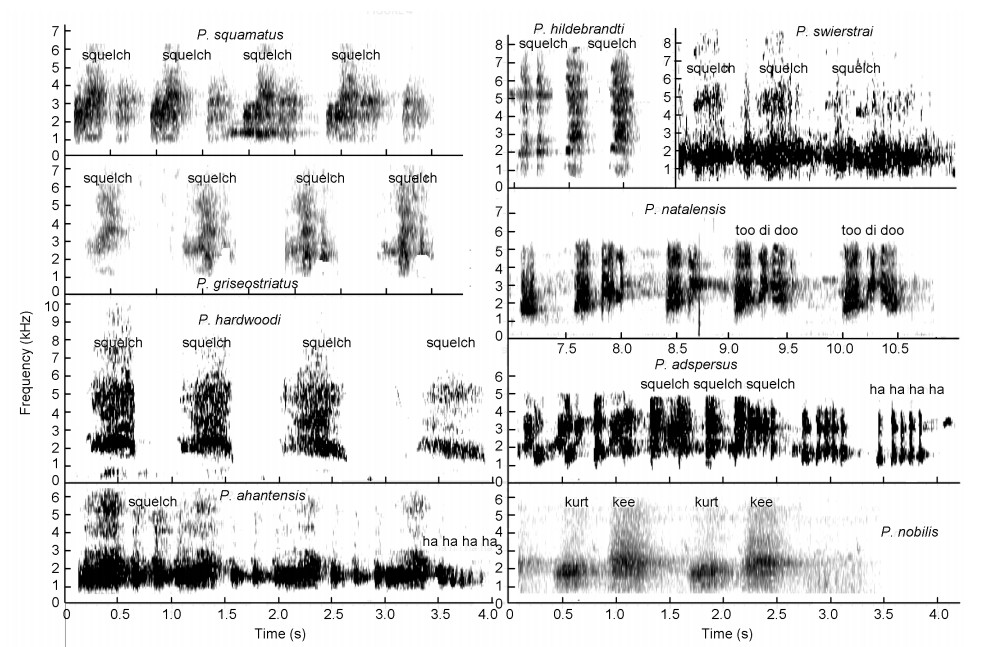
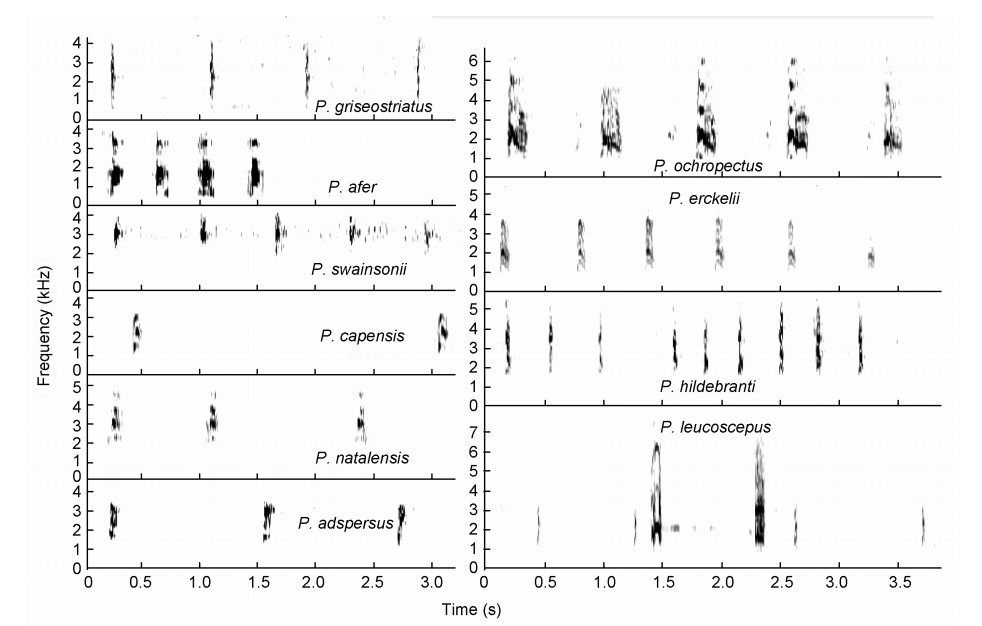
Squelching crows
These spurfowls (Fig. 4, Tables 3) have a much-accentuated loud sharp squelching crow without a pronounced upsweep shape in the syllable as is often the case with the guttural or low-intensity guttural crows. Five sound groups emerged (Fig. 2):
Figure
4.
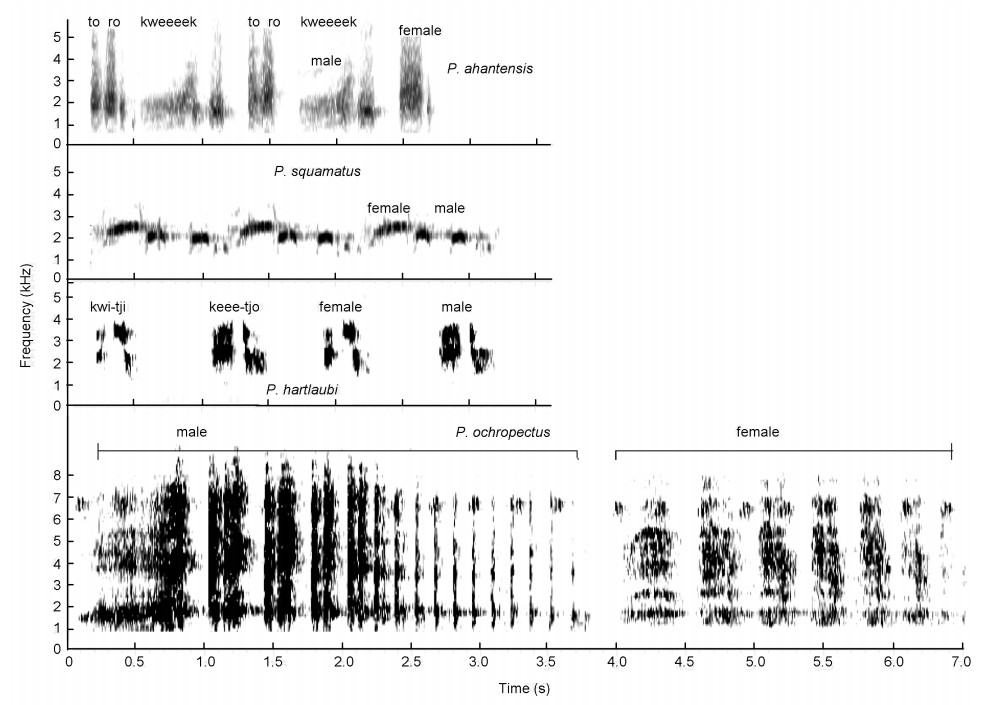
Squelching crows of Pternistis spurfowls visualized as spectrograms. Note the laughing syllables ("ha-ha-ha") in Red-billed Spurfowls (P. adspersus) but they do not dominate the call in terms of duration. The squelching crows of Ahanta (P. ahantensis) and Handsome (P. nobils) Spurfowls have two syllables hence the onomatopoeic labelling of the squelching notes ("kee" and "kweek").
(1) the Montane squelching group including the Grey-striped (P. griseostriatus) and Hardwood's (P.hardwoodi) Spurfowls that utter a series of monotonous, broad frequency bandwidth, squelching syllables, and Handsome (P. nobilis) and Swierstrai (P. swierstrai) Spurfowls that utter frequency-modulated, two-orthree syllable, squelch crows (Fig. 4);
(2) secondly, the chorus squelching group consisting of Hildebrandt (P. hildebrandti), Natal (P. natalensis) and Red-billed (P. adspersus) Spurfowls. Natal and Hildebrandt's Spurfowls are sister species and were retained as such (Bloomer and Crowe, 1998). Among this sound group a number of close-by conspecific males invariably respond, which results in a chorus sound with multiple short squelching syllables, or otherwise also described as cacophonies. These calls were often repeated continuously for 20–30 min just before roosting by Natal and Red-billed Spurfowls.;
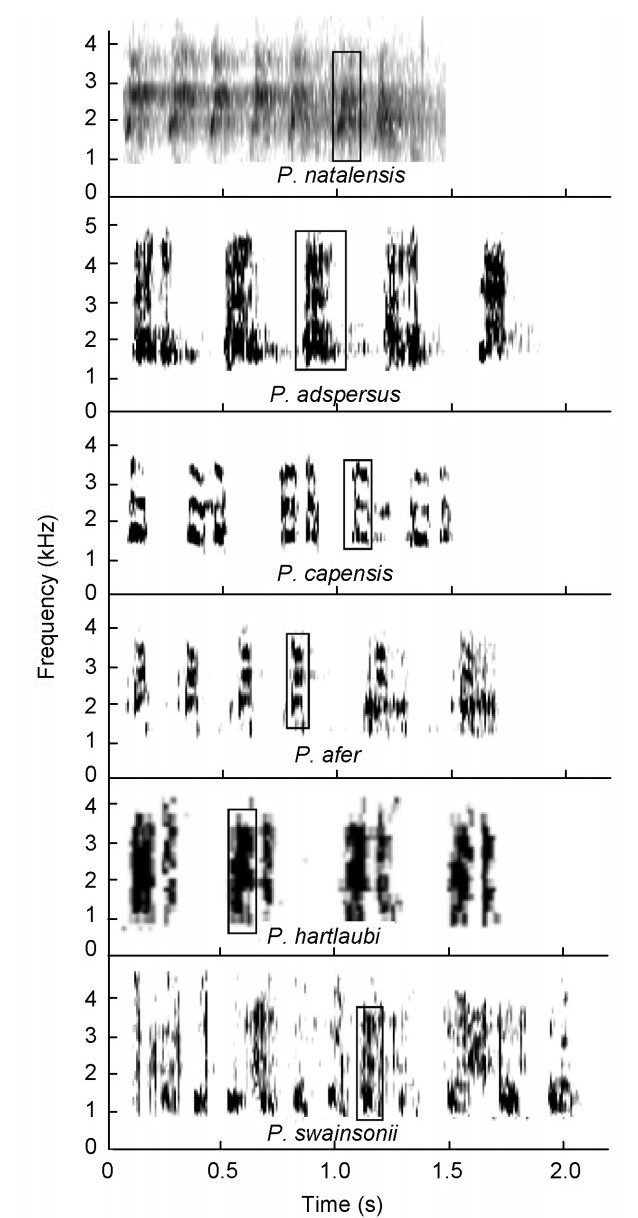
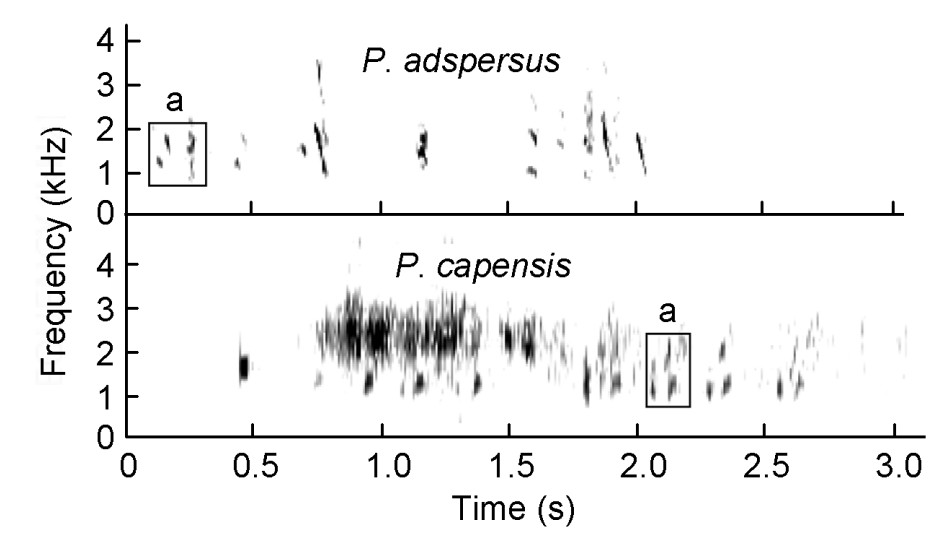
(3) the scaly tonal group consisting of Scaly (P. squamatus) and Ahanta (P. ahantensis) Spurfowls which have the squelching syllable, and in addition both utter duets with tonal syllables and the Scaly Spurfowl possesses an advertisement call with tonal syllables (Table 3, Figs. 5 and 6);
Figure
5.
Duets by Pternistis spurfowls visualized as spectrograms
Figure
6.
Advertisement calls of Pternistis spurfowls without crows visualized as spectrograms with tonal qualities. a = lower frequency syllables than crows with narrow frequency bandwidth. Note the horizontally oriented structure of the notes which is a solid texture and not perforated as it is with crows.
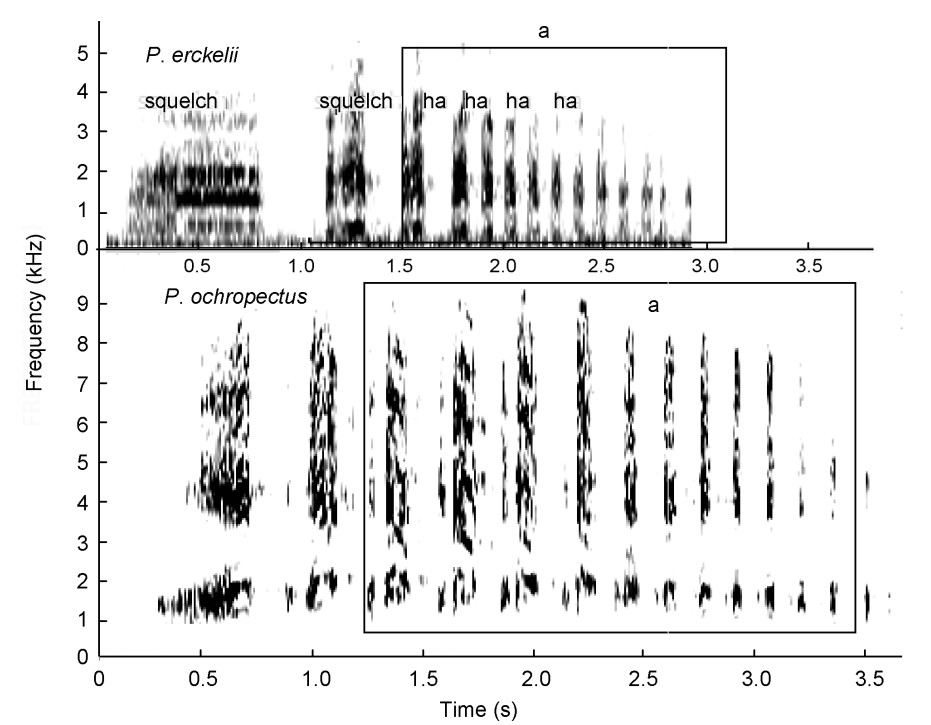
(4) the Montane laughing group consists of Djibouti and Erckel's Spurfowls (Fig. 7, Table 3). Their calls start with a squelching crow but most of the call is dominated (duration) with a series of clearly uttered, single staccato-type laughter notes. These two spurfowls are closest relatives and were retained as such (Madge and McGowan, 2002) and;
Figure
7.
High-intensity squelching crows with laughter syllables uttered by Erckel's (P. erckelii) and Djibouti (P. ochropectus) Spurfowls visualized as spectrograms. a = dominating laughter syllables in both calls. Both calls start with a squelching syllable.
(5) the Hartlaub's Spurfowl squelching duet group with only the one species as the name indicates (Fig. 5). This species is unique in the sense that it is the only spurfowl that utters a duet crow without a male-only crow. Furthermore, Tables 4 and 5 show the call repertoires and similarity coefficient (2 X calls shared by two species/total calls of both species) between all spurfowl species in southern Africa based on sound characteristics (also see Table 3 for differences with spurfowls in the rest of Africa). The Hartlaub's Spurfowl is the least similar to any other spurfowl (comparing similarity coefficients between Hartlaub's and the rest of the spurfowls, ANOVA: F = 120.05, df1,13, p < 0.01). The similarity coefficient between all spurfowls, setting aside Hartlaub's Spurfowl, varies from 0.56–0.82 (χ2 = 13.8, df1,14, p > 0.05) whereas the similarity coefficient between Hartlaub's Spurfowl and the rest varies from 0.13–0.18. Tonal-guttural crows
Table
4.
Arrangement of the southern Africa spurfowls Pternistis according to similar sound characteristics
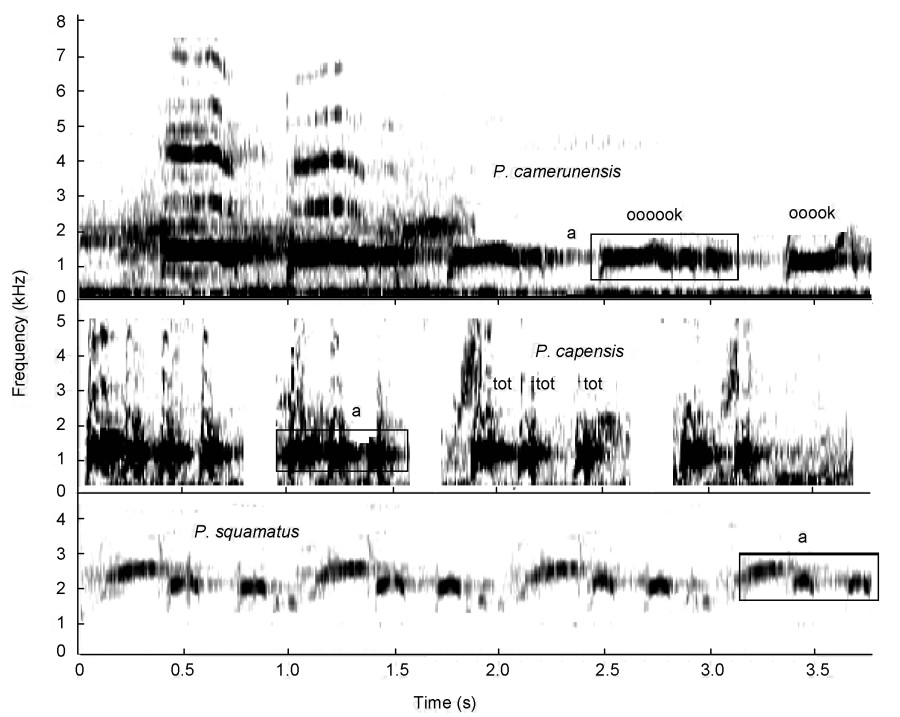
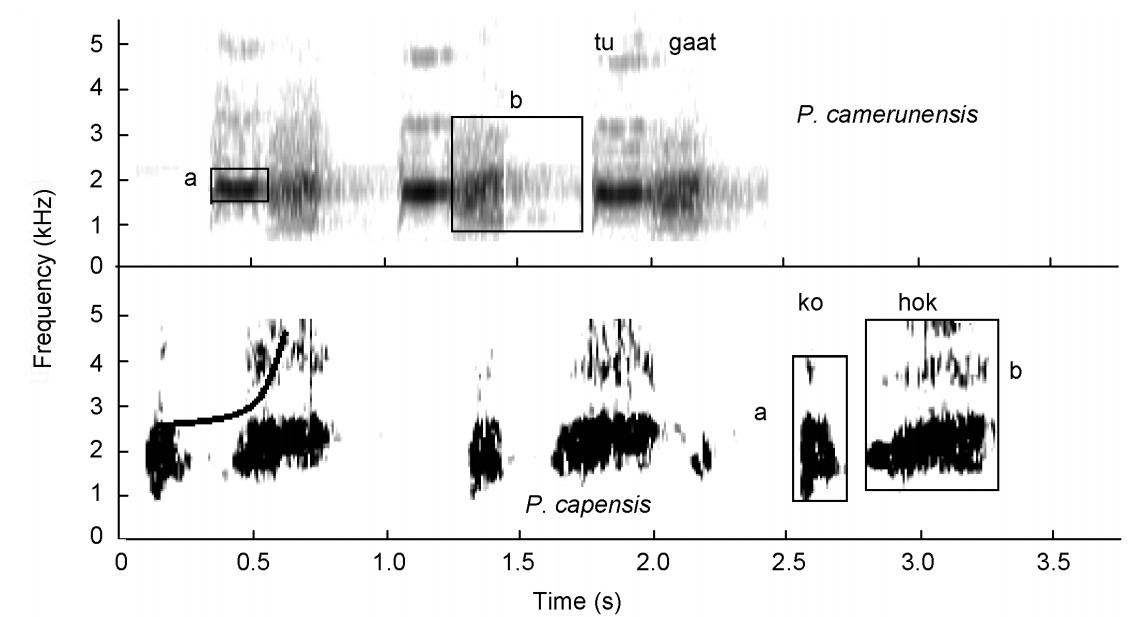
One sound group, the tonal-guttural crow group (Fig. 2), was formed that includes Cape and Mount Cameroon Spurfowls. These loud crows retain the basic shape of a crow, such as uttered by the bare-throated spurfowls, with an upsweep shape (Fig. 8), but they contain clear tonal elements mixed with guttural elements (Tables 3). However, Cape Spurfowl also uttered this call without any guttural elements (n = 3). On a secondary level, these species also share tonal advertisement calls without crow elements (Table 3).
Figure
8.
Tonal-guttural calls by Cape (P. capensis) and Cameroon (P. camerunensis) Spurfowls visualized as spectrograms. Note upsweep shape of syllable and the two elements (selection boxes) of the upsweep syllable in P. capensis. a = tonal element of syllable (narrower frequency bandwidth), b = guttural element of syllable (vertically oriented with a perforated-smudged texture).
These low-intensity, medium-loud (but often low-volume) crows have broad frequency bandwidths, strong upsweep syllable shapes and a strong propensity toward harmonics on the spectrogram (Fig. 9, Tables 2 and 3). Low-intensity guttural crows allowed minimal leverage for partitioning among species due to their homologous structure throughout the genus. This low-intensity guttural crows differed significantly in terms of spectrogram measurements compared to high-intensity guttural crows (t = 2.34, df1,9 = 75, p = 0.033, Table 2). Among the southern African spurfowls these calls were uttered by males, mainly when they detected females nearby during breeding; say 20–30 m away. This call was often uttered by the male chasing the female and was a prelude to courtship behavior.
Figure
9.
Low-intensity guttural crows of Pternistis spurfowls visualized as spectrograms. Note the homologous upsweep shape throughout the genus.
As expected, it was not possible to delimitate species, sound or species groups within the scope of maintenance calls as there is no profoundly intrinsic logic behind why these calls would have been subjected to sexual selection pressures like male advertisement calls (cf. Newton, 2003).
Distress calls
Distress calls consist of two variations of the same call and both are loud calls (Figs. 10 and 11). Based on the selected spectrogram measurements (see Table 2) the difference between the alarm calls on the ground and the flight alarm calls was found to be non-significant (t = 1.268, df = 7, p = 0.245). However, flight calls were uttered at a faster mean rate of 4.1 syllables/second (n = 5, SD = 1.38, range = 2.5–5.5) producing a "kwara-kwara-kwara" sound effect and ground calls were slower at 1.47 syllables/second (n = 9, SD = 0.72, range = 0.57–2.6) (ANOVA: F = 22.67, df1,12, p < 0.01).
Figure
10.
Alarm calls of Pternistis spurfowls on the ground visualized as spectrograms
Figure
11.
Flight alarm calls of Pternistis spurfowls during flight visualized as spectrograms. The Hartlaub's Spurfowl's (P. hartlaubi) spectrogram was abstracted from Maclean (1985). The square block indicates two harmonics for each syllable above the fundamental frequency (base frequency below harmonics).
The alarm call on the ground was a high-volume call with a broad frequency bandwidth that produces harmonics, but with a very short syllable (Table 2 and Fig. 10). This homologous call was recorded in at least 15 spurfowl species across Africa. It is a cluck ("chitchit") call often heard when the bird encounters an intruder such as a dog, human, rabbit or mongoose nearby. Some species, such as Red-billed and Natal Spurfowls, varied the number of clucks per call but the sound was otherwise homologous.
The flight alarm call was a high-volume call with a broad frequency bandwidth that produces harmonics (Fig. 11 and Tables 2 and 3). Flight calls were uttered when the bird was disturbed on the ground to a point where it fled, making the call during flight.
Cheep calls by chicks
The cheep call was a medium-loud short cheep with a typical arrow-head shape when visualized as a spectrogram (Fig. 12, Table 3). These chick calls were low volume, with a narrow frequency bandwidth with no harmonics. The cheeps were homologous for all spurfowl species recorded in South Africa and also recorded in Ahanta and Djibouti Spurfowls and are expected to be representative of most spurfowls in the rest of Africa. It is unknown if they are used by Hartlaub's Spurfowl. The cheeps were typically uttered when the mother and chicks were on the move and the chicks became temporarily isolated from the covey. In all southern African spurfowls, except Hartlaub's Spurfowl, the mother is the sole caretaker of her offspring (Komen, 1987; van Niekerk et al., 2009; J.H. van Niekerk, unpublished).
Figure
12.
Calls (cheeps) of Pternistis spurfowl chicks visualized as spectrograms. Note the homologous arrow-head-shaped syllables.
The medium-loud mew (whining) call is homologous throughout the genus that allowed no partitioning (Fig. 13). This call was found in all the spurfowls of southern Africa but also in spurfowl species in the rest of Africa, such as Djibouti and Yellow-necked Spurfowls (see Table 3). This call was of medium volume, narrow frequency bandwidth usually without harmonics with long syllables. It was uttered by sub-adults that had become detached from the family covey, say temporarily by about 3–5 m, and had been rejected by their mothers, or by sub-adults that were pecked by an adult male (also see van Niekerk, 1983).
Figure
13.
Juvenile calls (whining "mews") of Pternistis Spurfowls visualized as spectrograms
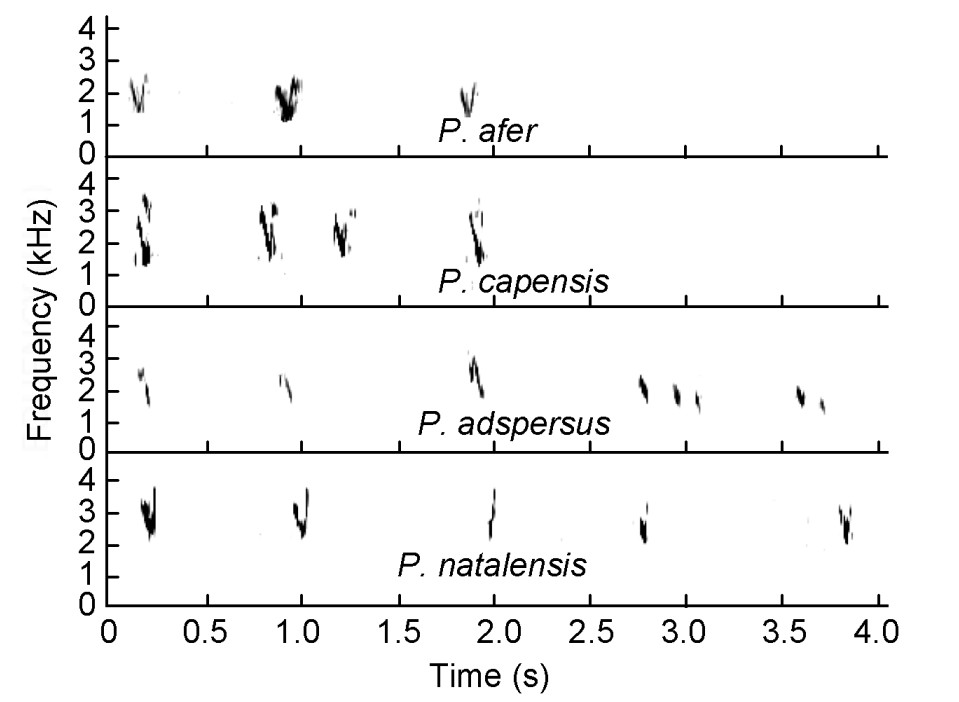
Cape, Natal and Red-billed Spurfowls uttered soft, short-syllabled feeding calls during foraging (Fig. 14). These were often uttered by Cape Spurfowl males (n = 5) that foraged alone, about 30–50 m away from any other conspecific, meaning that they do not possess any social significance.
Figure
14.
Feeding calls of Pternistis spurfowls visualized as spectrograms. a = similar syllables/elements in both species.
These calls consist of male-female and mother-offspring bonding calls. They consist mainly of medium-loud or soft short syllables which set them apart from the loud advertisement calls.
Mother-offspring bonding calls
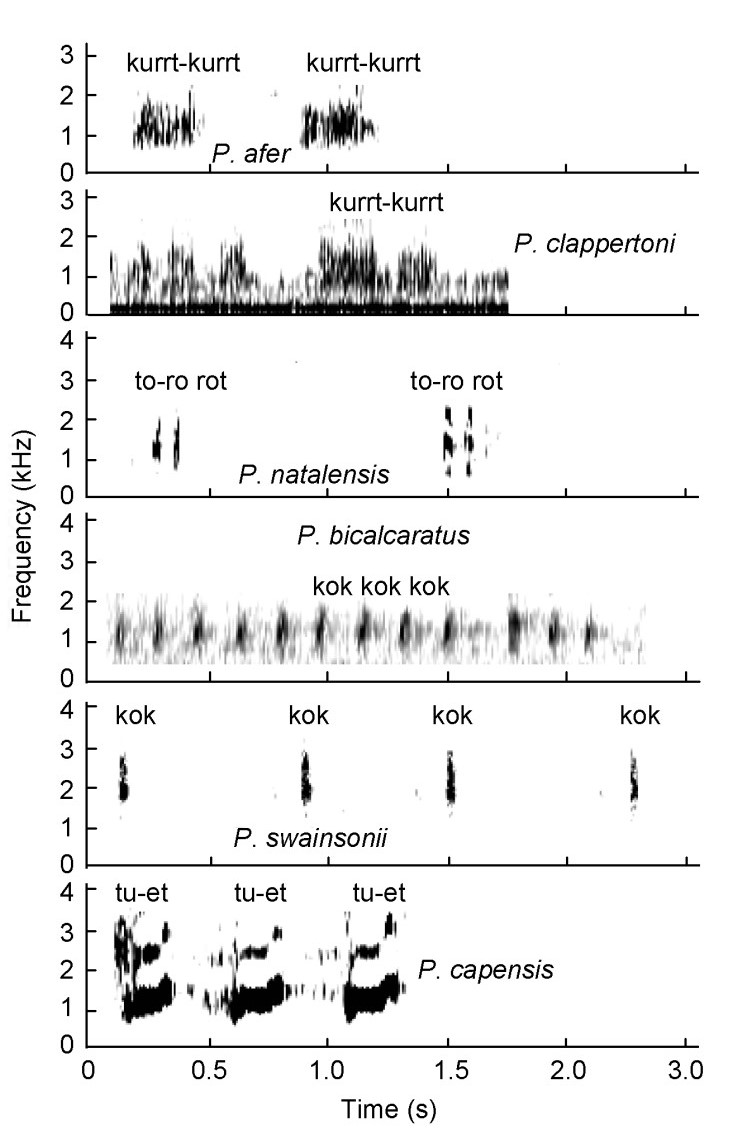
Mother-offspring bonding calls were uttered by the mother when intruders such as humans or dogs approached the nursing covey and are medium-loud calls. These calls by different species differed quite substantially from one another in terms of spectrogram shape and measurements (Fig. 15). In Red-necked Spurfowl (guttural grunting "kurrt-kurrt") and Cape Spurfowl ("tu-et tu-et tu-et") the syllable was longer than in Natal ("to-ro-rot") and Swainson's Spurfowl ("kok kok kok"). The call of Cape Spurfowls was of a higher volume compared with the other species. The guttural grunting call of Red-necked Spurfowls was also recorded for Clapperton's (P. clappertoni) and Doublespurred Spurfowls. Similarly, the "kok kok kok" sounds by Swainson's Spurfowl were also recorded for Doublespurred (n = 2) and Grey-striped (n = 2) (P. griseostriatus) Spurfowls. The "to-ro rot" sound of Natal Spurfowls was also recorded for Scaly (n = 3) and Red-billed Spurfowls (n = 3).
Figure
15.
Mother-offspring bonding calls of Pternistis spurfowls visualized as spectrograms. Note spectrogram shape differences between species.
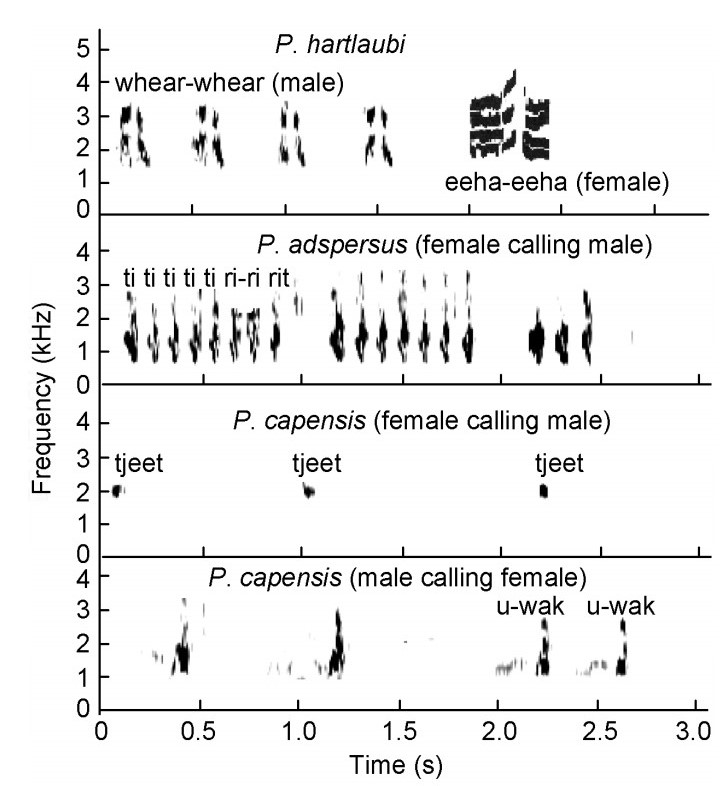
These calls are medium to loud. Female Red-billed Spurfowls gave a "ti ti ti ti ti – ri rit" call when separated from the male (Fig. 16). The male responded with its advertisement call. Male and female Cape Spurfowls gave "u-wak u-wak" and "tjeet tjeet" calls respectively when temporarily separated by vegetation over a distance of say 5–15 m. Hartlaub's Spurfowl males uttered a "whear-whear" and females responded with an "eehaa-eehaa" call during courtship but the male's call was also uttered when a playback was conducted showing its discontent with intruders. When the Red-billed Spurfowl female call was played back to a Cape Spurfowl male, on five occasions it approached and responded vocally each time with an advertisement call.
Figure
16.
Male-female bonding calls by Pternistis spurfowls in southern Africa visualized as spectrograms (female Hartlaub's Spurfowl's (P. hartlaubi) call abstracted from Komen, 1987)
Of all the species tested in southern Africa, only Swainson's and Natal Spurfowls gave no vocal response to their own advertisement calls (Table 6).
Table
6.
Responses to advertisement playback calls by Pternistis spurfowls in southern Africa. The response was either an advertisement call or a duet by P. hartlaubi.
The advertisement calls of all the southern Africa spurfowls were played to each other (n = 3) and no interspecific response (vocal or courtship display) was elicited.
Intra-specific mew (whining) call tests
Cape (n = 5) and Red-necked (n = 4) Spurfowl's offspring responded to their own mew calls, but not Natal Spurfowl (n = 5). Swainson's and Hartlaub's Spurfowls were not tested. In response to the mew call of the Natal Spurfowl, the mother Cape Spurfowl responded with a mother-offspring bonding call (n = 3).
Inter-specific mew call tests
When the "mew" call of Cape Spurfowl was played back to Red-necked (n = 3), Red-billed (n = 3) and Swainson's Spurfowls (n = 1), the nearby males approached their females and began courtship displays by standing in an upright posture near the female with body feathers erect, as observed in all southern African species except Hartlaub's Spurfowl for which the test was not conducted (van Niekerk, 1983). However, when the "mew" call of the Cape Spurfowl was played back to the Natal Spurfowl (n = 5), there was no display or vocal response. This display was also elicited from a Cape Spurfowl male when a Natal Spurfowl mew was played back to it (n = 4).
Intra-specific alarm call tests
When the ground alarm call (Fig. 10) was played back to Swainson's (n = 5) and Natal Spurfowl (n = 3) no response was elicited. The other species were not tested.
Inter-specific alarm call tests
The ground alarm calls of Cape Spurfowl did not entice any response from Natal (n = 3) and Swainson's Spurfowl (n = 5). However, when the ground alarm call of Natal Spurfowl was played back to Cape Spurfowl, three mother-chick coveys responded by running for nearby cover, while the rest, including single territorial males (n = 8) and sub-adult coveys that were without their parents (n = 3), did not respond.
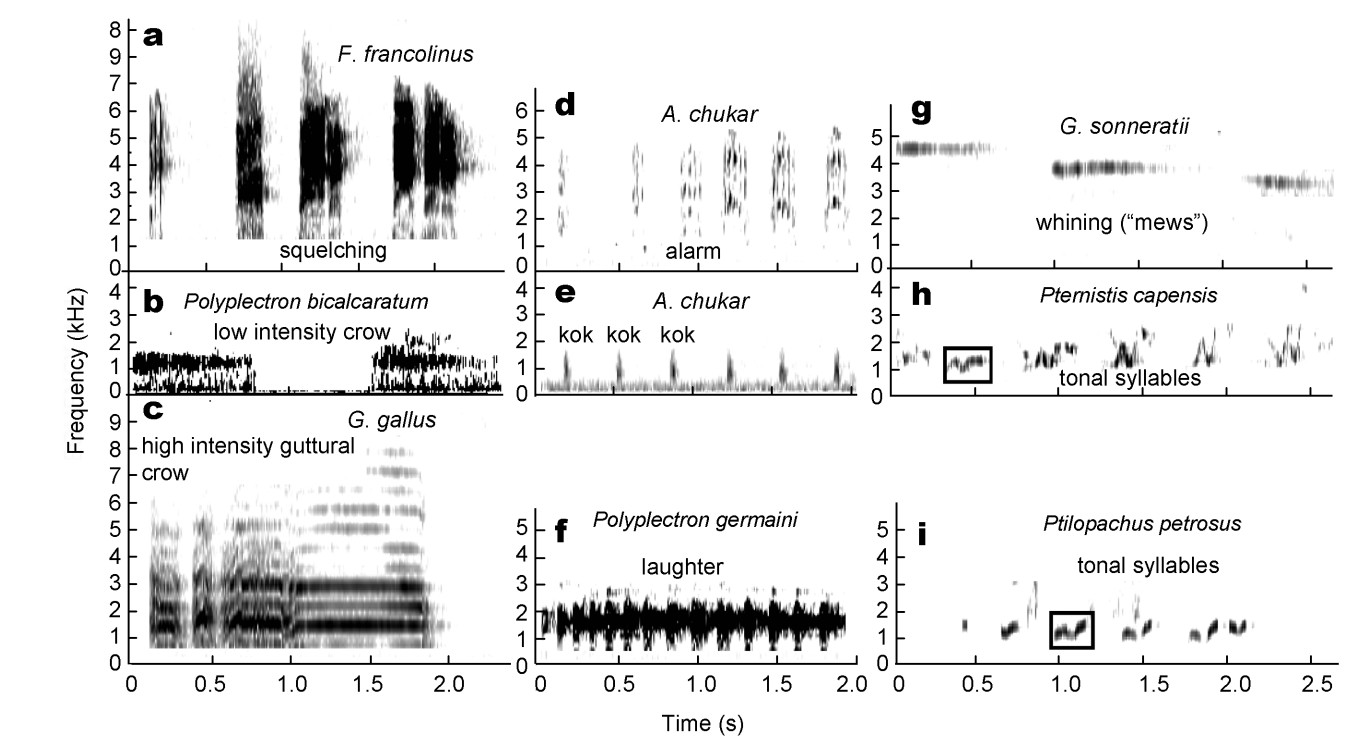
Table
7.Pternistis spurfowl sounds and syllables identified in a selection of outgroup species in the family Phasianidae (Order: Galliformes)
Sound/syllable
Outgroup species that uttered the sounds/syllables found in Pternistis spurfowls
High intensity crows
Red Junglefowl (Gallus galluss), Grey Junglefowl (G. sonneratii), Green Junglefowl (G. varius), Chukar Partridge (Alectorus chukar), Germain's Peacock-pheasant (Polyplectron germaini)
Low intensity crow alarm calls
Grey Peacock-pheasant (P. bicalcaratum), Red Junglefowl, Domestic Chicken (G. domesticus), Swamp Francolin, Chinese Francolin (F. pintandeanus), Grey Francolin (F. pondicerianus), Black Francolin (F. francolinus), Harlequin Quail (Corturnix delegorguei), Chinese Bamboo-partridge (Bambusicola thoracica)
Whining ("mews") "kok kok" call
Germain's Peacock-pheasant, Ring-necked Pheasant (Phasianus colchicus), Grey Junglefowl, Green Junglefowl, Chukar Partridge, Great Argus (Argusianus argus), Tibetan Snowcock (Tetraogalus tibetanus)
Laughter syllable
Germain's Peacock-pheasant, Chukar Partridge
Guttural syllable
Chukar Partridge, Red Junglefowl, Ring-necked Pheasant, Dometsic Chicken, Germain's Peacock Pheasant, Grey Junglefowl, Chinese Francolin
Tonal sound
Stone Partridge (Ptilopachus petrosus), Nahan's Partridge (P. nahani)
Figure
17.
Visualized spectrograms of calls from selected outgroup species of the family Phasianidae (Order: Galliformes) that are compared to the calls of Pternistis spurfowls. The tonal notes of Cape Spurfowl (P. capensis) were included to show homologous structures of syllables.
In all the southern African species as well as the Djibouti Spurfowl, advertisement calls are territorial calls. These calls are also expected to be territorial calls for species occurring in the rest of Africa given the obvious structural similarities of calls, but behavioral studies for these species were not detailed enough to state this categorically (Madge and McGowan, 2002).
Unlike Swainson's and Red-necked Spurfowls (the bare-throated guttural group) that occupy relatively large territories, members of the chorus squelching group tend to behave more compactly, i.e. males are closer and are especially heard before or after roosting in tree thickets that occur in pockets near water (and seasonal streams) whereas Swainson's and Red-necked Spurfowls do not roost in trees and males do not live close by each other.
Generally, duets are regarded as advertisement calls as they most certainly are a mechanism used by both males and females to demarcate their position in a territory or a breeding site. They may also enable breeding pairs to establish the location of neighbouring coveys with whom their offspring can mate since siblings do not pair (e.g. Komen, 1987; van Niekerk, 2009, 2010, 2012; J.H. van Niekerk, unpublished; Potts, 2012).
The tonal-guttural calls with tonal elements made by Cape and Cameroon Spurfowls are probably a variation of the crow ("kowaak") typically uttered by the barethroated spurfowls (Fig. 8). The basic structure of their calls is similar (Figs. 3 and 8). Both possess relatively drawn-out syllables ending with an upsweep element (a rise in frequency). Cape and Mount Cameroon Spurfowls do not form a conclusive sound group. Mount Cameroon Spurfowls live in a forest which explains the presence of tonal sounds in their repertoire since lowfrequency tonal sounds are less distorted in thick vegetation (Catchpole and Slater, 2008). These tonal syllables have been identified in the outgroup (Fig. 17) and therefore their occurrence in the Cape Spurfowl was not surprising but possibly a reversal. This resurgence might have been prompted by the dense fynbos habitat structure in which it lives. Among the southern African spurfowls low-intensity guttural crows were uttered by males, mainly when they detected females nearby during breeding; say 20–30 m away. It is expected that this behavior will be found in most other spurfowl species, as the call was also detected in Scaly, Doublespurred, Hardwood and Yellow-necked Spurfowls. The difference between the high-intensity guttural crow and low-intensity guttural crows in Pternistis spp., for those species that produce both, is probably caused by the difference in air pressure (volume) over the syrinx, but the social significance of the different outcomes is important. The loud advertisement crow must be able to cover the male's territorial/advertisement area maximally. On the other hand, the low-intensity guttural crow is uttered by males advertising their presence to nearby females. Cape and Natal Spurfowls do not possess a loud-intensity guttural crow but only a low-intensity guttural crow. It corroborates the purpose of the low-intensity guttural crow as a medium-loud call to attract females in the territory and not to maximize territorial size.
The flight alarm call was also found to be more or less similar to the flight calls of Helmeted Guineafowl (Numida meleagris), Crested (D. sephaena) and Orange River (S. lavaillantoides) Francolins (van Niekerk, 2010, 2012; J.H. van Niekerk, unpublished). The flight call of Hartlaub's Spurfowl is similar to the rest of the spurfowls as indicated by a spectrogram produced in the past (Maclean, 1985). However, Komen (1987) also recorded a flight alarm call with wing beats similar to the Scleroptila francolins (also see van Niekerk, 2010, 2012).
Cape, Natal and Red-billed Spurfowls uttered soft, short-syllabled feeding calls during foraging (Fig. 14). These calls are expected to occur in more Pternistis spurfowls. The divergence of mother offspring bonding call may serve to maximize the extinction of hybrid offspring (Newton, 2003).
The reluctance of Swainson's Spurfowl to respond to advertisement playback calls is possibly a ploy to allow younger males to enter after which they are repelled with brutal physical clashes. This may in fact also lure young females to the territorial males since young females are often associated with young males (cf. van Niekerk, 1983, 2002). Older males crow daily from separate territories while younger males often wonder around. In the case of Natal Spurfowl adult males often roost nearby each other and the crows of different males might be a strategy to distract/confuse predators at roosts, hence they do not respond but tolerate it (J.H. van Niekerk, unpublished). The low intensity crow might be a more important mechanism to attract females during courtship.
The fact that Swainson's and Natal Spurfowls did not respond to intra-specific alarm calls can possibly be regarded as a natural tendency by spurfowls not to reveal their respective positions to potential predators. The fact that Cape Spurfowls responded to inter-specific alarms from Natal Spurfowls showed that mothers with small chicks are highly tuned in to sounds in the immediate environment in order to keep her offspring aware of potential intruders such as humans, birds of prey or carnivores.
Although basic syllables such as the guttural, squelch, laughter and tonal sounds were combined in different sequences by different spurfowls to utter different calls (e.g. Red-billed Spurfowl uttered a guttural combined with laughter syllable and Cape Spurfowl uttered a tonal combined with a guttural syllable as in Table 3), I still could not infer derived characteristics in a coherent manner (also see Deregnaucourt, 2010). In fact, the mixing of sound syllables/elements to produce different calls is common in birds (Catchpole and Slater, 2008).
Congruence between sound groups and existing monophyletic groups
Table 3 shows the resultant sound groups arranged against the monophyletic groups of Hall (1963). There was substantial congruence between the sound groups and the monophyletic groups by Hall (Table 3). Hall's groups were also supported by Bloomer and Crowe (1998) who added a DNA component, in the sense that selected members of her groups were examined and fitted into the separate monophyletic groups (Table 3). However, Madge and McGowan (2002) and Hall (1963) grouped Hartlaub's Spurfowl with the southern vermiculated group. Notwithstanding that there were squelching syllables in the Hartlaub's Spurfowl's advertisement duet calls, it was placed in its own sound group. However, due to its significant dissimilarity with Pternistis calls in general (Tables 5 and 6), it was marked with a question mark in Table 3, and therefore is regarded as a preliminary designation awaiting more study. Bloomer and Crowe (1998) also described the Hartlaub's Spurfowl's relationship as uncertain. Bloomer and Crowe (1998) found a close relationship between Double-spurred and Scaly Spurfowls but both Madge and McGowan (2002) and the present study found them to be home in the vermiculated and scaly groups respectively. Madge and McGowan (2002) treated Hardwood's Spurfowl as being a member of the northern vermiculated group. This present study found its sound to be homologous with the sound structures of the Montane group.
Finally, both Bloomer and Crowe (1998) and Madge and McGowan (2002) grouped the Cape Spurfowl with the southern vermiculated group. Due to the overwhelming tonal syllables in its call it was placed in a separate tentative sound group which is not part of the vermiculated group (Fig. 2). However, from a morphobehavioral and DNA point of view, it fitted into the southern vermiculated group (Bloomer and Crowe, 1998). Therefore, despite its tonal sounds, the preferred view that is maintained is that the Cape Spurfowl belongs to the southern vermiculated group. In other words, the sound similarities between Cape and Cameroon Spurfowls are regarded as a matter of ecological convergence (Newton, 2003).
Divergence of advertisement calls
Advertisement calls is a category in which substantial divergence occurred between species (or species groups) when compared to maintenance calls. At least 13 different advertisement calls were identified (Table 3) compared to no partitioning within the mew, cheep and alarm calls. During sound-playbacks southern African spurfowls responded vigorously to their own advertisement calls (Table 6) but none responded to the loud advertisement calls of other spurfowls, suggesting that the advertisement call is an important inter-specific buffer to prevent cross-breeding. Sound discrimination is deemed especially important as spurfowls possess very few secondary sexual characteristics (e.g. bright feathers or prominent crests) that can be used to attract females (cf. Deregnaucourt, 2010; Kimball et al., 2011).
Male-female bonding calls (Fig. 16) also have a reproductive purpose and as expected differed substantially from species to species. Likewise, mother-offspring bonding calls also differed to some extent between species suggesting that mothers "fingerprinted" this call as a security mechanism to invest in their own progeny during nurturing. Although too few samples were obtained (i.e. not a representative from each of Hall's group) to make inferences across the genus, it nevertheless suggests that these bonding sounds probably also diverged to minimize hybridization and deserve future study.
Juxtaposing the loud advertisement crows which have wide frequency bandwidths (with a propensity toward harmonics), the softer "mew" call, with its narrower frequency bandwidth (less harmonics), remained remarkably stable throughout Pternistis (Fig. 13). In fact, Cape and Red-billed Spurfowls responded to "mews" from other spurfowl species when they were played back to them. This call remained stable as there was, relatively speaking, little selective social pressure to change as it is a call mainly uttered within the context of the family/sibling group (between sub-adults and mothers), where it serves no reproductive purpose. It is used over short distances of about 20–30 m as opposed to loud territorial crows that have to reach distances of 100–150 m (suggested by territorial sizes, e.g. van Niekerk, 2004, 2009, 2011, and unpublished).
Mew (whining) calls are mostly associated with subadults that are separated from their mothers, but also have the effect of enticing territorial males to court nearby females, even if the mew call is uttered by subadults (or by sound-playback). This is a surprise since Cape, Natal and Red-necked Spurfowl males pecked at sub-adults that utter this sound (van Niekerk, 1983). The possible reason for territorial males responding sexually to mew calls is that the mew calls of offspring broadcast that adult females are nearby and on arrival males have to compete with juveniles/sub-adults for the mother's/female's attention. Indeed, Swainson's and Hartlaub's Spurfowl females utter the mew call similar to the sub-adult mew call, during courtship (Komen, 1987).
The alarm call on the ground and during flight respectively also showed strong homology throughout the genus (Figs. 10 and 11, and Table 2). These loud calls have short syllables and frequency bandwidths with harmonics. Unlike advertisement calls between adult spurfowls, these calls are not bonding calls between males and females, nor are they calls that are used between territorial rivals during fitness battles (e.g. van Niekerk et al., 2009), but rather they are used to drive birds into cover away from intruders. As these calls have by nature no reproductive purpose, there have probably been few evolutionary pressures to diverge discernibly.
Despite the dissimilarity between Hartlaub's Spurfowl calls on the one hand and the rest of the spurfowls on the other hand (Tables 4 and 5), this spurfowl remains related to the genus (Pternistis) based on (1) its squelch crow sound and; (2) its mew call that is also uttered by juveniles and adult Hartlaub's Spurfowl females, as is the case with Swainson's Spurfowl females. Its flight call is also similar to other Pternistis spp. but this call's structure is common in the outgroup as well (Fig. 17).
Conclusions
The repertoire size of spurfowls varies from 7–11 calls. One must be mindful that as more field work is conducted more call types could be unravelled. The common occurrence of the guttural sound (high- and low-intensity guttural crows) and the whining call ("mew") of juveniles is absent in all the francolins in Africa. Therefore, the evidence is in favour of the fact that spurfowls (Pternistis) and francolins in Africa (Dendroperdix,Scleroptila, Peliperdix) form two clades, i.e. not a monophyletic entity.
My sound groups suggest an early speciation from one ancestor into at least guttural and squelching groups that are further basal than the monophyletic groups of Hall (1963). Hartlaub's Spurfowl could have been an early offshoot of the "squelchers".
This evidence suggests that loud advertisement calls that are used for reproduction purposes have undergone substantial changes that are probably intended to minimize hybridization. The arrangement of spurfowl species into "natural sound groups" provided an opportunity to reassess the phylogeny of spurfowls from a sound perspective that has hitherto not been done.
Although it was not possible to determine polarity in spurfowl sounds intrinsically, it was possible to align different sound groups in a congruent manner with the existing monophyletic spurfowl groups proposed by Hall (1963). Encouragingly, this paper demonstrated that sound groups did not split previously recognized most closely-related species including (1) Swainson's and Red-necked Spurfowls (Little and Crowe, 2011), (2) Ecrkell's and Djibouti Spurfowls (Madge and McGowan, 2002) and (3) Natal and Hildebrandt's Spurfowls (Little and Crowe, 2011) (see Fig. 2).
Sound credits
Credits for calls and sounds assessed from www.xenocanto.org: G. Wagner XC2775 (Ahanta Spurfowl); L. Hansen XC28978, XC28979, XC28980 (Scaly Spurfowl); L. Boon XC41203 (Handsome Spurfowl); D. Moyer XC83807, XC83835 (Heuglin's and Hildebrandt's Spurfowls); T. Spanhove XC57559, XC57554 (Harwood's Spurfowl); J. Poelstra XC83661 and B Piot XC66313 (Erckel's Spurfowl); M. Nelson XC9910, XC99150, XC99151, XC99152 (Double-spurred Spurfowl); D. Farrow XC37441 and R. O'Donnell XC91226 (Chukar Partridge); S. Bot XC44885 and D. Farrow XC37514 (Grey Junglefowl); D. Edwards XC70624 (Green Junglefowl); F. Lambert XC11108 (Tibetan Snowcock); C. Robson XC19258 (Germain's Peacock-pheasant); G. Wagner XC42527 (Grey Peacock-pheasant); F. Lambert XC58891 (Malayan Peacock-pheasant); C. Robson XC1917 (Great Argus); S. Bot XC22710 (Common Quail); D. Solomon XC45651 (Harlequin Quail).
Acknowledgements
Thank you to Klarissa van Niekerk, AnnElizabeth Jones, Melissa van Niekerk, Johann van Niekerk (junior) and Hansie Erasmus for assisting me during 10 years of field recordings throughout southern Africa and to the personnel of the Cornell Laboratory of Ornithology (Ithaca, New York) for training.
Catchpole CK, Slater PJB. 2008. Bird Song: Biological Themes and Variations. Cambridge University Press, Cambridge.
Charif RA, Waack AM, Strickman LM. 2010. Raven Pro 1.4 User's Manual. Cornell Laboratory of Ornithology, Ithaca, New York.
Crowe T. 2010. Phylogenetic affinities of enigmatic African galliforms: the Stone Partridge Ptilopachus petrosus and Latham's and Nahan's Francolins Francolinus lathami and F. nahani. Cladistics, 26: 206.
Crowe TM, Harley EH, Jakutowicz MB, Komen J, Crowe AA. 1992. Phylogenetic, taxonomic and biogeographical implications of genetic, morphological, and behavioural variation in francolins (Phasianidae: Francolinus). Auk, 109: 24–42.
Deregnaucourt S. 2010. Interspecific hybridization as a tool to understand vocal divergence: the example of crowing in quail (Genus Corturnix). PLoS ONE, 5(2): e9451.
Erbe C, King AR. 2008. Automatic detection of marine mammals using information entropy. J Acoust Soc Am, 124: 2833–2840.
Fleischer RC, Rothstein SI. 1988. Known secondary contact and rapid gene flow among subspecies and dialects in the Brownheaded Cowbird. Evolution, 42: 1146–1158.
Fuller RA, Akitw P, Amuno JB, Fuller CL, Ofwono JM, Proaktor G, Ssemmanda R. 2012. Using playback of vocalisations to survey the Nahan's Francolin, a threatened African forest galliform. Ostrich, 83(1): 1–6.
Hall BP. 1963. The francolins, a study in speciation. Bull Br Mus, 10: 8–204.
Harris MP. 1970. Abnormal migration and hybridisation of Larusargentatus and L. fuscus after interspecies fostering experiments. Ibis, 112: 488–498.
Hockey PAR, Dean WRJ, Ryan PG. 2005. Roberts Birds of southern Africa, VIIth ed. The Trustees of the John Voelcker Bird Book Fund, Cape Town.
Kimball RT, Colette M, Braun EL. 2011. A macroevolutionary perspective on multiple sexual traits in the Phasianidae (Galliformes). Int J Evol Biol, doi: .
Komen J. 1987. Preliminary observations of the social pattern, behaviour and vocalisations of Hartlaub's Francolin. S Afr J Wildl Res, 1: 82–86.
Little R, Crowe TM. 2011. Gamebirds of Southern Africa. Struik, Cape Town.
Little RM, Crowe TM. 1992. Vocal behaviour of Greywing Francolin Francolinus africanus can be used to estimate population density. Ostrich, 63: 172–179.
Maclean GL. 1985. Roberts Birds of Southern Africa. John Voelcker Bird Book Fund, Cape Town.
Madge S, Mcgowan P. 2002. Pheasants, Partridges and Grouse. Christopher Helm, London.
Mandiwana-neudani TG, Kopuchain C, Louw G, Crowe TM. 2011. A study of gross morphological and histological syringeal features of true francolins (Galliformes: Francolinus,Scleroptila, Peliperdix and Dendroperdix spp. ) and spurfowls (Pternistis spp. ) in a phylogenetic context. Ostrich, 82: 115–126.
Mills MSL. 2007. Vocalizations of Angolan Birds. Volume 1. Birds Angola and Birding Africa, Cape Town.
Milstein PLES, Wolff SW. 1987. The over-simplification of our francolins. S Afr J Wildl Res, 1: 58–65.
Newton I. 2003. The Speciation and Biogeography of Birds. Academic Press, Amsterdam.
Potts GR. 2012. Partridges. Harper Collins, London, UK.
Seddon N, Merrill RM, Tobias JA. 2008. Sexually selected traits predict patterns of species richness in a diverse clade of suboscine birds. J Evol Biol, 22: 623–636.
Seddon N, Tobias JA. 2007. Song divergence at the edge of Amazonia: an empirical test of the peripatric speciation model. Biol J Linn Soc, 90: 173–188.
Seddon N. 2005. Ecological adaptation and species recognition drives local evolution in neotropical suboscine birds. Evolution, 59: 200–215.
Sinclair I, RyanP. 2003. Birds of Africa South of the Sahara. Struik Publishers, Cape Town.
van Niekerk JH, Barendse M, Mare F. 2009. Behaviour of Rednecked Spurfowl Pternistis afer in the Boknes and Cannon Rock coastal resorts, Alexandria district, Eastern Cape province, South Africa. Ostrich, 80: 43–45.
van Niekerk JH. 1983. Observations on courtship in Swainson's Francolin. Bokmakierie, 35: 90–92.
van Niekerk JH. 2002. Notes on territoriality and pair-formation in two juvenile Swianson's Francolin. Ostrich, 73: 59–78
van Niekerk JH. 2004. Ecology and dispersion of Swainson's Spurfowl Pternistis swainsonii, introduced to a modified habitat in the Heidelberg district, Gauteng province, South Africa. Ostrich, 75: 178–180.
van Niekerk JH. 2009. Hybridization between Red-billed Pternistis adsperus and Natal P. natalensis Spurfowls in captivity calls for a search in the wild. Ostrich, 80: 63–64.
van Niekerk JH. 2010. Vocal behaviour of Crested Francolin Dendroperdix sephaena in response to playback calls. Ostrich, 81: 149–154.
van Niekerk JH. 2011. Habitat-use and range contraction of Swainson's Spurfowl at the Krugersdorp Game Reserve, Gauteng province, South Africa. Ostrich, 82: 43–47.
van Niekerk JH. 2012. Vocal behaviour of Orange River Francolin Scleroptila levaillantoides based on visual and sound playback surveys. Ostrich, 83: 147–152.
Viljoen PJ. 2005. AGRED'S Gamebirds of South Africa. African Gamebird Research Education and Development Trust, Johannesburg.
Wolff SW. 1978. The use of tape-recorder calls to locate and census Orange River Partridges. S Afr J Wildl Res, 8: 135–136.
Table
1.
List with Pternistis spurfowls and calls assessed, source of information and time spent observing southern African spurfowls species during fieldwork
Species
Number of recordings assessed and time spent observing southern African spurfowls
Source (and locality coordinates in the case of southern African spurfowls)
Table
3.
Calls of Pternistis spurfowls based on homologous sound principles (spectrogram structure) arranged in sound groups which were also compared with existing phylogenetic results for congruence/incongruence. Numbers between parentheses indicate sample sizes.
"??" = tentatively placed and "?" = further field work required.
Table
6.
Responses to advertisement playback calls by Pternistis spurfowls in southern Africa. The response was either an advertisement call or a duet by P. hartlaubi.
Table
7.Pternistis spurfowl sounds and syllables identified in a selection of outgroup species in the family Phasianidae (Order: Galliformes)
Sound/syllable
Outgroup species that uttered the sounds/syllables found in Pternistis spurfowls
High intensity crows
Red Junglefowl (Gallus galluss), Grey Junglefowl (G. sonneratii), Green Junglefowl (G. varius), Chukar Partridge (Alectorus chukar), Germain's Peacock-pheasant (Polyplectron germaini)
Low intensity crow alarm calls
Grey Peacock-pheasant (P. bicalcaratum), Red Junglefowl, Domestic Chicken (G. domesticus), Swamp Francolin, Chinese Francolin (F. pintandeanus), Grey Francolin (F. pondicerianus), Black Francolin (F. francolinus), Harlequin Quail (Corturnix delegorguei), Chinese Bamboo-partridge (Bambusicola thoracica)
Whining ("mews") "kok kok" call
Germain's Peacock-pheasant, Ring-necked Pheasant (Phasianus colchicus), Grey Junglefowl, Green Junglefowl, Chukar Partridge, Great Argus (Argusianus argus), Tibetan Snowcock (Tetraogalus tibetanus)
Laughter syllable
Germain's Peacock-pheasant, Chukar Partridge
Guttural syllable
Chukar Partridge, Red Junglefowl, Ring-necked Pheasant, Dometsic Chicken, Germain's Peacock Pheasant, Grey Junglefowl, Chinese Francolin
Tonal sound
Stone Partridge (Ptilopachus petrosus), Nahan's Partridge (P. nahani)

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds