Rose THOROGOOD, Nicholas B. DAVIES. 2013: Hawk mimicry and the evolution of polymorphic cuckoos. Avian Research, 4(1): 39-50. DOI: 10.5122/cbirds.2013.0002
Citation:
Rose THOROGOOD, Nicholas B. DAVIES. 2013: Hawk mimicry and the evolution of polymorphic cuckoos. Avian Research, 4(1): 39-50. DOI: 10.5122/cbirds.2013.0002
Rose THOROGOOD, Nicholas B. DAVIES. 2013: Hawk mimicry and the evolution of polymorphic cuckoos. Avian Research, 4(1): 39-50. DOI: 10.5122/cbirds.2013.0002
Citation:
Rose THOROGOOD, Nicholas B. DAVIES. 2013: Hawk mimicry and the evolution of polymorphic cuckoos. Avian Research, 4(1): 39-50. DOI: 10.5122/cbirds.2013.0002
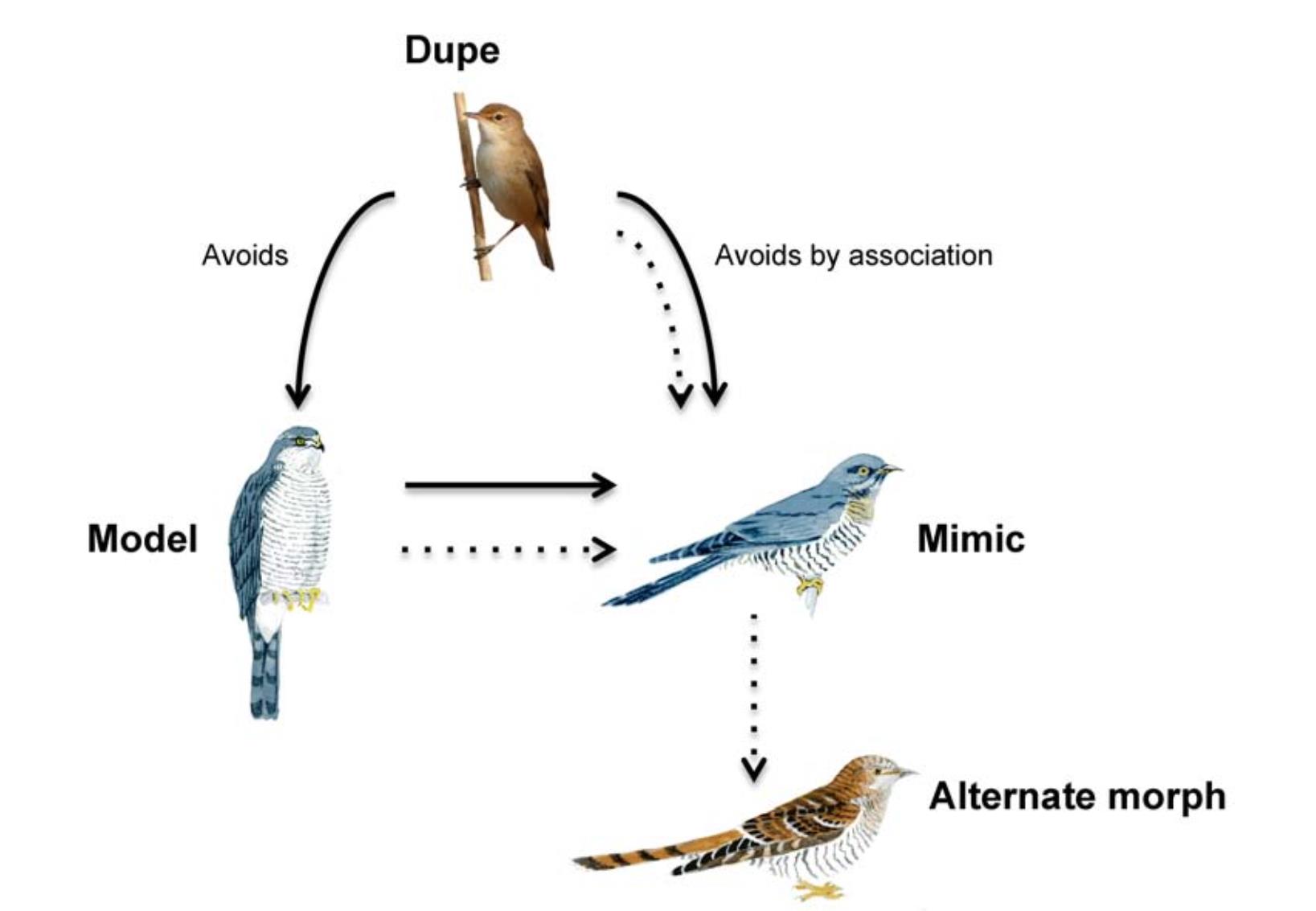
The resemblance of some parasitic cuckoos to Accipiter hawks has been known since ancient times. Recent experiments show that the hawk-like features of Common Cuckoos (Cuculus canorus) facilitate access to Reed Warbler (Acrocephalus scirpaceus) host nests. However, social information alerts hosts to see through the cuckoo's mimetic disguise. In turn, this has promoted the evolution of a cuckoo polymorphism to thwart host recognition. Here we show by comparative analyses that parasitic cuckoos with hawk-like features (yellow eyes, barred underparts, yellow legs) are more likely to be polymorphic (29% of species) than those without (8% of species). Phylogenetic analyses confirm correlated evolution of hawk-like features and cuckoo polymorphism. We suggest that mimicry dynamics are particularly likely to promote the evolution of various guises in parasitic cuckoos to beat host defences.
Endemic species have long been a key focus in conservation efforts (Myers et al., 2000), given that the level of endemics might be positively correlated with species richness (Lamoreux et al., 2006). Besides, endemic species with limited dispersal capacity might be sensitive to changes in local climate, or vulnerable to invasive species (Ohlemüller et al., 2008). A good understanding of evolutionary processes such as population subdivisions, changes of effective population size and genetic connectivity of endemic species would shed light on evolutionary processes as well as on conservation management.
The Blue Eared Pheasant (Crossoptilon auritum), belonging to Phasianidae, Galliformes, is a rare and endemic pheasant species in western China (Lei and Lu, 2006). Its wild populations are found at Helan Mountain, as well as along the eastern edge of the Qinghai-Tibetan Plateau (QTP), covering Qinghai, Gansu and Sichuan provinces (Lei and Lu, 2006). Although previous studies have been carried out on the biology and ecology of C. auritum (Sun et al., 2005; Li et al., 2009; Wu and Liu, 2010), a thorough assessment of genetic diversity is urgently needed to assess the population viability of this species. Furthermore, apart from C. auritum, the genus Crossoptilon includes three other species, i.e., C. mantchuricum, C. harmani and C. crossoptilon. These species are endemic to China (Zheng, 2011) and all are listed on the IUCN Red List of Threatened Species (IUCN 2011) because of a rapid decline in the size of their population, caused by habitat fragmentation and hunting (Lei and Lu, 2006). Given these concerns, obtaining molecular markers is a prerequisite in understanding the genetic background of C. auritum and might be useful for population genetic studies in Crossoptilon species.
Microsatellites are powerful tools for conservation genetic studies such as population genetics, mating systems and investigations into kinship (Primmer et al., 2005; Karl et al., 2011). Compared with isolated novel microsatellite markers, cross-species microsatellite amplification from closely related species is cost-effective (Zane et al., 2002). More importantly, it has been suggested that this method has successfully worked among species belonging to the same genera, different genera and even different families (Barbará et al., 2007; Huang and Liao, 2010). Given that numerous microsatellite markers have been developed for various Phasianidae species (Cheng et al., 1995; Wang et al., 2009; Zhou and Zhang, 2009), we attempted to establish microsatellite markers for C. auritum through cross-species amplification from a large number of marker candidates. In order to test the effectiveness of these markers, we also carried out preliminary parentage analysis among captive individuals of known pedigree.
Materials and methods
A total of 20 C. auritum blood samples from brachial veins were collected to develop microsatellite loci by cross-species amplification, nine of which were from the Linxia Zoo in Gansu Province and the other 11 from Huzhu in Qinghai Province, China. Additionally, nine individual birds from two families of the Beijing Wildlife Park in Daxing and Beijing Zoo were used to conduct parentage analysis. We have detailed the known pedigree of these two families of birds.
Genomic DNA was extracted using DNA extraction kits (Tian Gen Biotech, Beijing, China). The cross-species microsatellite markers (Table 1) came from various Galliforme species, including 16 loci from Meleagris gallopavo (Burt et al., 2003), 30 from Tragopan temminckii (Zhou and Zhang, 2009), 20 from Syrmaticus reevesii (Wang et al., 2009), 41 from Gallus gallus (Cheng et al., 1995; Dawson et al., 2010), four from C. mantchuricum (Zhao et al., Beijing Normal University, unpublished results) and one from Syrmaticus mikado (S.H. Li, Taiwan Normal University, unpublished results). After filtering out those loci with poor cross-amplification, the polymorphism of the remaining pairs were tested with either 6-FAM or HEX fluorescent dyelabeled on 5′ of a single forward primer. Polymerase chain reaction (PCR) was carried out in a 10 µL reaction system containing 100 ng DNA, 0.25 µL of each primer, 1 µL of a 10 × PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTP mix and 0.75 U Taq polymerase (Takara, Japan). The reaction was denatured at 94℃ for 5 min, followed by 40 cycles at 94℃ for 30 s, a touch-down annealing process from 58–47℃, reducing in steps of 0.5℃ per cycle and another 20 cycles annealing at 47℃ and then 72℃ for 50 s, with a final extension at 72℃ for 5 min. Fragment analysis was conducted on an ABI PRISM 3100 Genetic Analyzer using the GeneMapper software (Applied Biosystems) with ROX-500 as the standard for size. We conducted PCR amplification and fragment analysis at least twice to ensure the accuracy of individual genotypes. Sequencing of selected homozygotes was also conducted in both directions with amplification primers (BGI Bio Tech, Beijing, China) to ensure that the products were genuine microsatellites.
Table
1.
Characterization of 11 microsatellite loci
Locus
Primer sequences (5′–3′)
Accession No.
Repeat motifs
n
NA
Size range (bp)
HO
HE
Species origin
Chromosome
2580
F: TTAACCTATCAGGTCGTTGCG
AL592580
(CA)n
20
8
191–213
1.00
0.80
M. gallopavo
21
R: CAGTGCACATGCAGGCAG
3D2
F: TCTCTGACGTATCGCATCT
FJ221373
(GT)n
19
6
286–304
0.47
0.58
S. reevesii
4
R: ACTTCCCCTGGTAAACT
1H4
F: TGAACAAGTGAGGCGGAGC
/
(TG)n
20
10
127–161
0.65
0.81
S. reevesii
/
R: CTGCACACAGCCCGAAGC
2420
F: CATCATCTGCCAATGCAGAGG
/
(TTTA)n
20
4
118–142
0.55
0.54
S. mikado
/
R: AAGCCCATATATGCTTCCTGG
4H1
F: TATGAAACAGACTTAATCC
FJ221388
(GTTT)n
20
4
203–211
0.85
0.67
S. reevesii
1
R: TGCAGCATTTGAGTAAC
5C9
F: TATGGGAAATGTGTACCTTTA
GQ184557
(CA)n
20
10
221–259
0.95
0.89
C. mantchuricum
10
R: TCCAGGCAACACGTAACA
TT06
F: TGAGAGATTTTGACCCA
GQ181183
(CA)n
20
7
225–237
0.85
0.83
T. temminckii
6
R: CAAGACTTCACCCTACAGATA
4F8
F: GTGGCATGCCTAGTAGATGTT
/
(AC)n
20
11
186–214
0.75
0.88
C. mantchuricum
/
R: CCCTGTGGTACGAACTGTC
SR11
F: ATCAATATGGACTGCTCCGT
FJ221381
(TG)n
20
5
210–248
0.55
0.58
S. reevesii
17
R: TCCTTCAAGGCCAAGTG
5H7
F: CCAAGAGGGAGGCACACGTTC
U60782
(TG)n
20
3
186–194
0.55
0.42
G. gallus
8
R: AGCCATAAATAAGCAAACGC
4C12
F: ATAGGCGGACAGAGGATAGA
FJ221385
(CA)n
20
4
160–170
0.30
0.59
S. reevesii
Z
R: CCCCGCATCGAGGTG
Notes: n is the number of successfully genotyped individuals and NA the number of alleles. HO is observed heterozygosity and HE expected heterozygosity.
We blast the acquired microsatellite sequences from C. auritum on the chicken genome in GenBank to find their locations and to investigate the potential linkages among the loci. If some loci were mapped in sex chromosome, we carried out sexing identification with our sample set by using primer sex1/sex2 (Wang and Zhang, 2009) to cross-validate the sex-linkage loci.
We tested observed heterozygosity (HO), expected heterozygosity (HE), the number of allele per locus, the Hardy-Weinberg equilibrium (HWE) and linkage-disequilibrium in each population using Arlequinv.3.11 (Excoffier et al., 2005). We calculated the frequencies of null alleles by using FreeNA (Chapuis and Estoup, 2007). High polymorphic markers with HWE were used for paternity tests in CERVUS 3.0 with 100000 times simulations to estimate their resolving power. Significant levels were recorded after application of the sequential Bonferroni correction (Rice, 1989; Excoffier et al., 2005).
Results
Sixty-two (55.4%) of the 112 cross-species markers could be efficiently amplified in C. auritum, while 11 (17.7%) of the 62 markers had a moderate to high level of polymorphism (3–11 loci), with the expected heterozygosities ranging from 0.42 to 0.89 (Table 1). Locus 4C12 was found to be homozygous in heterogametic females and heterozygous in 56.2% of the males (n = 28) and thus most likely to link with the Z chromosome. Three loci (1H4, 2420 and 4F8) were not targeted on the chicken genome. All loci were at the HWE; neither linkage disequilibrium among pairs of loci nor null alleles was found. The parentage analysis results showed that if neither parent was known, the successful assignment rates were 98% at a 95% confidence level and 98% at an 80% confidence level. Paternity test results are consistent with known pedigrees.
Discussion
The phylogenetic relationship between the original and target species seems to affect the success of cross-species amplification (Primmer et al., 1996). In our analyses, cross-species amplification from Crossoptilon was the most effective (2/4 = 50%), followed by those from Syrmaticus (6/21 = 28.6%), Meleagris (1/16 = 6.25%), Tragopan (1/30 = 3.0%) and Gallus (1/41 = 2.4%). Phylogenetic analysis showed a successive relationship between Crossoptilon and the genera mentioned earlier (Kimball et al., 2011), indicating that evolutionary relationships may play an important role in cross-species amplification of microsatellite markers within Galliformes.
Although large numbers of microsatellite markers are found in autosomes, Z-linked microsatellites markers are still rarely available, even in the well-characterized chicken genome (Groenen et al., 2000). One Z-linked marker TUT, originally isolated from the Tetrao urogallus (Segelbacher et al., 2000, Wang et al., 2011), failed to be amplified in chickens and might be specific for Tetraoninae grouse. In the present study, the Z-linked polymorphic locus 4C12 seems to have general application in different Galliforme species and might yet provide a valuable tool in kinship and demographic analyses combined with other loci.
Andersson M. 1995. Evolution of reversed sex roles, sexual size dimorphism, and mating system in coucals (Centropodidae, Aves). Biol J Linn Soc, 54: 173–181.
Bond AB. 2007. The evolution of color polymorphism: crypticity, searching images, and apostatic selection. Annu Rev Ecol Evol S, 38: 489–514.
Brooke M de L, Davies NB, Noble DG. 1998. Rapid decline of host defences in response to reduced cuckoo parasitism: behavioural flexibility of Reed Warblers in a changing world. P R Soc Lond B, 265: 1277–1282.
Burns K, Shultz A. 2012. Widespread cryptic dichromatism and ultraviolet reflectance in the largest radiation of Neotropical songbirds: implications of accounting for avian vision in the study of plumage evolution. Auk, 129: 211–221.
Campobello D, Sealy SG. 2011. Use of social over personal information enhances nest defense against avian brood parasitism. Behav Ecol, 22: 422–428.
Clarke BC. 1969. The evidence for apostatic selection. Heredity, 24: 347–352.
Davies NB, Brooke M de L. 1988. Cuckoos versus reed warblers: adaptations and counteradaptations. Anim Behav, 36: 262–284.
Davies NB, Welbergen JA. 2008. Cuckoo-hawk mimicry? An experimental test. P R Soc B, 275: 1817–1822.
Davies NB, Welbergen JA. 2009. Social transmission of a host defense against cuckoo parasitism. Science, 324: 1318–1320.
del Hoyo J, Elliott A, Sargatal J. 1994. Handbook of the Birds of the World, Vol. 2. New World Vultures to Guinea Fowl. Lynx Edition, Barcelona, Spain. Endler J. 1981. An overview of the relationships between mimicry and crypsis. Biol J Linn Soc, 16: 25–31.
Ferguson-Lees J, Christie DA. 2001. Raptors of the World. Christopher Helm, London, U.K.
Fowlie MK, Krüger O. 2003. The evolution of plumage polymorphism in birds of prey and owls: the apostatic selection hypothesis revisited. J Evolution Biol, 16: 577–583.
Galeotti P, Rubolini D, Dunn PO, Fasola M. 2003. Colour polymorphism in birds: causes and functions. J Evolution Biol, 16: 635–646.
Gärtner K. 1981. Das Wegnehmen von Wirtsvogeleiern durch den Kuckuck Cuculus canorus. Ornithologische Mitteilungen, 33: 115–131.
Gluckman T-L, Cardoso GC. 2010. The dual function of barred plumage in birds: camouflage and communication. J Evolution Biol, 23: 2501–2506.
Grafen A. 1989. The phylogenetic regression. Philos T Roy Soc B, 326: 119–157.
Grim T. 2005. Mimicry vs. similarity: which resemblances between brood parasites and their hosts are mimetic and which are not? Biol J Linn Soc, 84: 69–78.
Holen Ø, Johnstone R. 2004. The evolution of mimicry under constraints. Am Nat, 164: 598–613.
Honza M, Procházka P, Šicha V, Požgayová, M. 2010. Nest defence in a cuckoo host: great reed warblers risk themselves equally for their own and parasitic chicks. Behaviour, 147, 741–756.
Honza M, Šicha V, Procházka P, Ležalová R. 2006. Host nest defense against a color-dimorphic brood parasite: great reed warblers (Acrocephalus arundinaceus) versus Common Cuckoos (Cuculus canorus). J Ornithol, 147: 629–637.
Johnsgard PA. 1997. The Avian Brood Parasites: Deception at the Nest. Oxford University Press, Oxford, U.K.
Krüger O, Davies NB, Sorenson MD. 2007. The evolution of sexual dimorphism in parasitic cuckoos: sexual selection or coevolution? P R Soc B, 274: 1553–1560.
Kunte K. 2009. The diversity and evolution of batesian mimicry in Papilio swallowtail butterflies. Evolution, 63: 2707–2716.
Lai C. 1998. Messenger of Spring and morality: cuckoo lore in Chinese sources. J Am Oriental Soc, 118: 530–542.
Lyon BE, Gilbert GS. In press. Unparasitized species mob and alarm call to cuckoos: implications for sparrowhawk mimicry by brood parasitic cuckoos. Wilson J Ornithol.
Moksnes A, Røskaft E, Korsnes L. 1993. Rejection of cuckoo (Cuculus canorus) eggs by meadow pipits (Anthus pratensis). Behav Ecol, 4: 120–127.
Møller AP, Solonen T, Byholm P, Huhta E, Nielsen JT, Tornberg R. 2012. Spatial consistency in susceptibility of prey species to predation by two Accipiter hawks. J Avian Biol, 43: 390–396.
Newton I. 1986. The Sparrowhawk. T & AD Poyser, London, U.K.
Pagel M, Meade A. 2006. Bayesian analysis of correlated evolution of discrete characters by reversible-jump Markov chain Monte Carlo. Am Nat, 167: 808–825.
Pagel M. 1999. Inferring the historical patterns of biological evolution. Nature, 401: 877–884.
Pasteur G. 1982. A classification review of mimicry systems. Annu Rev Ecol Evol S, 13: 169–199.
Payne RB. 1967. Interspecific communication signals in parasitic birds. Am Nat, 101: 363–375.
Payne RB. 2005. The Cuckoos. Oxford University Press, Oxford, U.K.
Pough F. 1988. Mimicry of vertebrates: are the rules different? Am Nat, 131: S67–S102.
Roulin A. 2004. The evolution, maintenance and adaptive function of genetic colour polymorphism in birds. Biol Rev, 79: 815–848.
Ruxton GD, Sherratt TN, Speed MP. 2004. Avoiding Attack: the Evolutionary Ecology of Crypsis, Warning Signals and Mimicry. Oxford University Press, New York.
Seddon N, Tobias JA, Eaton M, Ödeen A, Byers BE. 2010. Human vision can provide a valid proxy for avian perception of sexual dichromatism. Auk, 127: 283–292.
Soler M, Soler J. 2000. Brood-parasite interactions between great spotted cuckoos and magpies: a model system for studying coevolutionary relationships. Oecologia, 125: 309–320.
Sorenson MD, Payne RB. 2005. A molecular genetic analysis of the cuckoo phylogeny. In: Payne RB (ed) The Cuckoos. Oxford University Press, Oxford, U.K. pp 68–94.
Stoddard MC. 2012. Mimicry and masquerade from the avian visual perspective. Curr Zool, 58: 630–648.
Thorogood R, Davies NB. 2012. Cuckoos combat socially transmitted defenses of reed warbler hosts with a plumage polymorphism. Science, 337: 578–580.
Trnka A, Prokop P, Grim T. 2012. Uncovering dangerous cheats: how do avian hosts recognize adult brood parasites? PloS ONE, 7: e37445.
Trnka A, Prokop P. 2012. The effectiveness of hawk mimicry in protecting cuckoos from aggressive hosts. Anim Behav, 83: 263–268.
Turner JRG. 1978. Why male butterflies are non-mimetic: natural selection, sexual selection, group selection, modification and sieving. Biol J Linn Soc, 10: 385–432.
Vane-Wright RI. 1979. Towards a theory of the evolution of butterfly colour patterns under directional and disruptive selection. Biol J Linn Soc, 11: 141–152.
Voipio P. 1953. The hepaticus variety and the juvenile plumage types of the cuckoo. Ornis Fennica, 30: 99–117.
Wallace AR. 1889. Darwinism: An Exposition of the Theory of Natural Selection with Some of Its Applications. Macmillan, London, U.K.
Welbergen JA, Davies NB. 2009. Strategic variation in mobbing as a front line of defense against brood parasitism. Curr Biol, 19: 235–240.
Welbergen JA, Davies NB. 2011. A parasite in wolf's clothing: hawk mimicry reduces mobbing of cuckoos by hosts. Behav Ecol, 22: 574–579.
Wickler W. 1968. Mimicry in Plants and Animals. World University Library, London, U.K.
Table
1.
Characterization of 11 microsatellite loci
Locus
Primer sequences (5′–3′)
Accession No.
Repeat motifs
n
NA
Size range (bp)
HO
HE
Species origin
Chromosome
2580
F: TTAACCTATCAGGTCGTTGCG
AL592580
(CA)n
20
8
191–213
1.00
0.80
M. gallopavo
21
R: CAGTGCACATGCAGGCAG
3D2
F: TCTCTGACGTATCGCATCT
FJ221373
(GT)n
19
6
286–304
0.47
0.58
S. reevesii
4
R: ACTTCCCCTGGTAAACT
1H4
F: TGAACAAGTGAGGCGGAGC
/
(TG)n
20
10
127–161
0.65
0.81
S. reevesii
/
R: CTGCACACAGCCCGAAGC
2420
F: CATCATCTGCCAATGCAGAGG
/
(TTTA)n
20
4
118–142
0.55
0.54
S. mikado
/
R: AAGCCCATATATGCTTCCTGG
4H1
F: TATGAAACAGACTTAATCC
FJ221388
(GTTT)n
20
4
203–211
0.85
0.67
S. reevesii
1
R: TGCAGCATTTGAGTAAC
5C9
F: TATGGGAAATGTGTACCTTTA
GQ184557
(CA)n
20
10
221–259
0.95
0.89
C. mantchuricum
10
R: TCCAGGCAACACGTAACA
TT06
F: TGAGAGATTTTGACCCA
GQ181183
(CA)n
20
7
225–237
0.85
0.83
T. temminckii
6
R: CAAGACTTCACCCTACAGATA
4F8
F: GTGGCATGCCTAGTAGATGTT
/
(AC)n
20
11
186–214
0.75
0.88
C. mantchuricum
/
R: CCCTGTGGTACGAACTGTC
SR11
F: ATCAATATGGACTGCTCCGT
FJ221381
(TG)n
20
5
210–248
0.55
0.58
S. reevesii
17
R: TCCTTCAAGGCCAAGTG
5H7
F: CCAAGAGGGAGGCACACGTTC
U60782
(TG)n
20
3
186–194
0.55
0.42
G. gallus
8
R: AGCCATAAATAAGCAAACGC
4C12
F: ATAGGCGGACAGAGGATAGA
FJ221385
(CA)n
20
4
160–170
0.30
0.59
S. reevesii
Z
R: CCCCGCATCGAGGTG
Notes: n is the number of successfully genotyped individuals and NA the number of alleles. HO is observed heterozygosity and HE expected heterozygosity.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds