Jin-Won LEE, Piotr G. JABŁOŃSKI. 2012: Egg color polymorphism and morph-ratio variation in Korean populations of the Vinous-throated Parrotbill. Avian Research, 3(4): 312-319. DOI: 10.5122/cbirds.2012.0036
Citation:
Jin-Won LEE, Piotr G. JABŁOŃSKI. 2012: Egg color polymorphism and morph-ratio variation in Korean populations of the Vinous-throated Parrotbill. Avian Research, 3(4): 312-319. DOI: 10.5122/cbirds.2012.0036
Jin-Won LEE, Piotr G. JABŁOŃSKI. 2012: Egg color polymorphism and morph-ratio variation in Korean populations of the Vinous-throated Parrotbill. Avian Research, 3(4): 312-319. DOI: 10.5122/cbirds.2012.0036
Citation:
Jin-Won LEE, Piotr G. JABŁOŃSKI. 2012: Egg color polymorphism and morph-ratio variation in Korean populations of the Vinous-throated Parrotbill. Avian Research, 3(4): 312-319. DOI: 10.5122/cbirds.2012.0036
Understanding the occurrence of multiple distinct phenotypes in a population of a species, i.e., polymorphism, is one of the challenges encountered in evolutionary biology. Egg color polymorphism in birds is one example of morphological polymorphism and disruptive selection has been proposed as a hypothetical mechanism to explain its occurrence. We studied how polymorphic egg colors (immaculate blue and white) occur in Korean populations of the Vinous-throated Parrotbill (Paradoxornis webbianus). Egg color ratios (the proportion of nests with blue eggs in a population) were monitored over a large spatial scale and egg colors were quantified using a spectrophotometer. We found egg color ratios to vary spatially among populations. Interestingly, there was a latitudinal morph-ratio cline in egg color ratios. The proportion of nests with blue eggs increased considerably with the latitude declined towards the southern part of the Korean peninsula. There were some quantitative variations in egg colors among populations. However, the pattern of variations was not consistent with those of the population egg color ratios. Based on these results, we discuss a potential scenario for the evolution of egg color polymorphism in the Vinous-throated Parrotbill.
In biology, polymorphism indicates that multiple discrete morphs co-occur in a population of a species with random mating (Ford, 1945; Roulin, 2004). Polymorphic species are widespread in nature, occurring across all life forms including plants and animals (Jones et al., 1977; Hoffman and Blouin, 2000; Gigord et al., 2001; Jonsson and Jonsson, 2001; Galeotti et al., 2003; John, 2003; Gray and McKinnon, 2007) and thus understanding the evolutionary forces that cause and maintain polymorphism is an important challenge in evolutionary biology and biodiversity. Avian egg polymorphism is one example of morphological polymorphism that can be easily studied in nature. Continuous variations among individuals in background color and spotting patterns of their eggs are found in many bird speices. However, some species, such as brood parasites and their hosts, often produce eggs with distinct colors and spotting patterns among individuals in a population (Collias, 1993; Kim et al., 1995; Ortega, 1998; Yang et al., 2010; Spottiswoode and Stevens, 2011). Although efforts have been made to understand the genetics, ecology and evolution of egg color polymorphism in this system of avian brood parasitism, our knowledge is still incomplete.
Takasu's (2003) analysis provided the theoretical background of the evolution of egg color polymorphism in the system of avian brood parasitism. He hypothesized mathematically that even if the initial hostparasite interaction starts with a continuous spectrum of egg appearances it could result in polymorphic egg phenotypes in both the parasite and the host as a result of disruptive selection. Some Paradoxornis species (e.g., the Ashy-throated Parrotbill (Paradoxornis alphonsianus), the Vinous-throated Parrotbill (P. webbianus)) and their brood parasite, the Common Cuckoo (Cuculus canorus) show clear egg color polymorphism representing one of best examples where this hypothesis could be tested. Yang et al. (2010) suggested disruptive selection as an evolutionary origin of egg color polymorphism observed in a system of the Common Cuckoo–Ashy-throated Parrotbill, in which both species lay polymorphic eggs with blue, white, and rarely pale blue coloration. They showed empirically that, similar to the Vinous-throated Parrotbill (Lee and Yoo, 2004), the host rejected parasitic eggs with different probabilities in proportion to the contrast in egg appearance between the host and parasite. They then suggested that the egg color ratio in the host population could be determined by natural selection in a frequency-dependent manner. Based on this finding, they hypothesized that the original morph was pale blue and that disruptive selection has evolved bluer and whiter eggs, resulting in egg color polymorphism. This hypothesis was later formalized mathematically by Liang et al. (2012) who determined how egg color polymorphism is maintained by selection in a system of Paradoxornis species and the Common Cuckoo. The model showed that frequencydependent selection may oscillate the morph ratio of egg phenotypes and that the pattern of oscillations may be affected by the inheritance mode of egg color, host sensitivity in detecting parasitic eggs and the selectivity of the parasite for host egg phenotypes.
Empirical data that can be used to test these theoretical predictions, and thus to elucidate the evolution of egg color polymorphism, are still lacking. Strictly speaking, the study of Yang et al. (2010) only confirmed the feasibility of frequency-dependent selection as a maintenance mechanism of egg color polymorphism rather than proving that disruptive selection is the selective force responsible for the occurrence and evolution of egg color diversification. In order to test this hypothesis properly, as Takasu (2005) suggested, we should address several issues. First, we should determine how egg color is inherited by daughters since the mode of inheritance mechanisms (asexually or sexually) could have different consequences of the arms race between the cuckoo and the parrotbill. Second, we need to quantify egg appearance objectively for a long period or across different populations in order to reveal any correlation between egg appearance and the intensity of brood parasitism. Finally, as Liang et al. (2012) suggested, in order to elucidate the origin and maintenance mechanism of egg color polymorphism in this system we need to monitor the morph ratio of egg phenotypes over a large spatialtemporal scale in both hosts and parasites.
Across a large spatial scale, we studied the egg color polymorphism in a Paradoxornis species, the Vinousthroated Parrotbill, a primary host of the Common Cuckoo in Korea. This species occurs from Mongolia, through China, Vietnam, Taiwan to Korea and the south-east Russian region of Ussuriland. However, their center of distribution is located in central China (Robson, 2007). In the Korean peninsula, they breed in open-cupped nests, lay clutches of 4–6 eggs and show egg color dimorphism: females either lay clutches of immaculate blue or white eggs (Kim et al., 1995). Lee (2008) reported that some individuals lay intermediate pale blue eggs but the frequency is negligible. It is already known that each female of this species has constant egg colors throughout her life (Kim et al., 1995), so the trait is highly likely to be determined genetically.
In our study, we addressed two key questions that are important for understanding the co-evolution between the Common Cuckoo and the Vinous-throated Parrotbill. First, does the morph ratio of host egg phenotypes (the proportion of nests with blue eggs) vary among populations? Second, does egg color (e.g., blue-green chroma) vary among populations in a manner predictable from the assumption that intense parasitism causes the evolution of polymorphic eggs? Based on previous studies (Takasu, 2005; Yang et al., 2010), we finally predict that egg colors should be bluer and/or whiter in populations with more white clutches than in populations with less white egg clutches.
Methods
Fieldwork
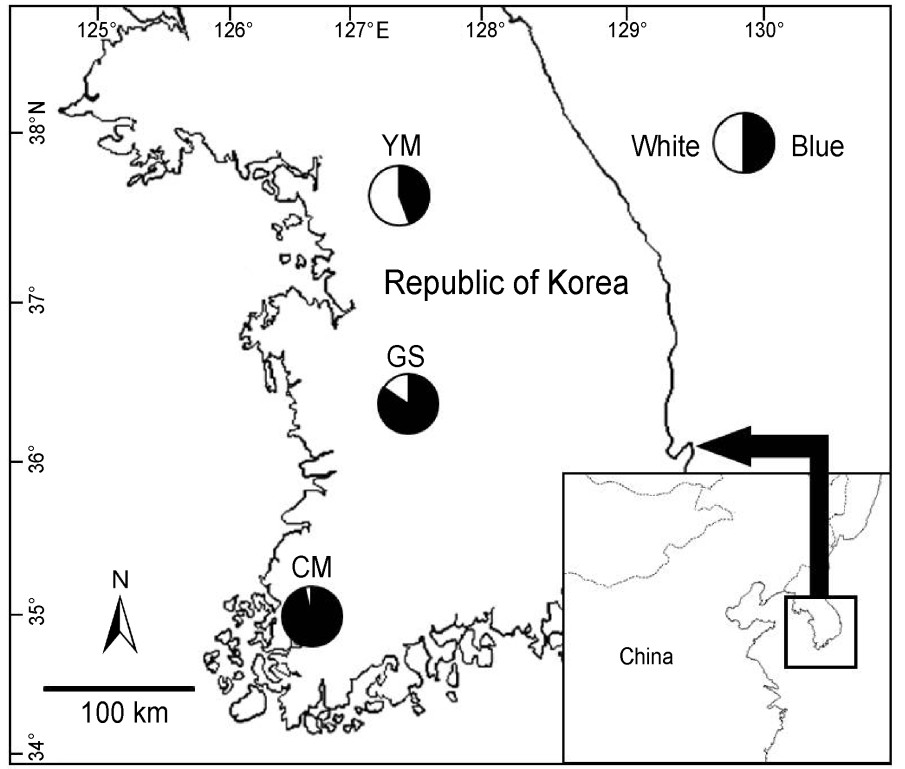
Fieldwork for determining egg color ratio and quantifying egg color was conducted in 2010 and 2011 at three geographically distant populations along the Korean peninsula: Yangseo-myeon (YM), Yangpyeonggun, Gyonggi-do (37°32'N, 127°20'E); Gyeryong-si (GS), Chungcheongnam-do (36°17'N, 127°14'E) and Cheonggye-myeon (CM), Muan-gun and Jeollanamdo (34°54'N, 126°26'E) (Fig. 1). We aimed to sample about 30 nests at each site where the egg color ratio of each population could be determined. Egg color was measured from the eggs of clutches around the onset of incubation (i.e., from 1 or 2 days before to 5 days after clutch completion). In total, we measured 42 blue eggs (8 clutches) and 16 white eggs (4 clutches) at GS, 78 blue eggs (17 clutches) and 4 white eggs (1 clutch) at CM and 56 blue eggs (12 clutches) and 69 white eggs (17 clutches) at YM (Table 1).
Figure
1.
Map of study area and morph-ratio cline. Circles represent relative proportion of blue and white egg nests in each location. Number of nests: YM = 29, GS = 32, CM = 25.
Reflectance spectra in the 300–700 nm range at 0.37 nm intervals were taken in the field using an Ocean Optics USB 2000+ spectrophotometer with a DT-Mini-2 light source and the Spectra Suite (Ocean Optics) software. Measurements were carried out using the probe (Ocean Optics reflection probe: QR230-7-XSR/BX) placed at a constant distance (ca. 1 mm in diameter of illuminated areas) from the surface of eggs at a 45° angle in a specially designed box with a probe holder (Ocean Optics RPH-1) which allowed us to remove the effect of ambient light. Reflectance spectra were calculated relative to a standard white reference (Labsphere USRS-99-010) and to complete darkness, which were calibrated prior to the measurement of each clutch. The egg of the Vinous-throated Parrotbill is immaculate and constant in color, so we measured color at three randomly selected points of each egg. There measurements were taken at each point, and the mean values used in the statistical analyses.
Blue-green chroma (BGC) was calculated as the proportion of total reflectance (R300–700) in the blue-green region (R400–550) (Moreno et al., 2006; Siefferman et al., 2006). The average BGC of all eggs in each clutch was used in the analysis. We also measured the consistency of BGC of each egg and among all eggs in each clutch by calculating the repeatability (r) of BGC between measurements within an egg and among eggs within a clutch using F statistics.
Statistics
We used one-way ANOVA to analyze the geographic variations among three populations in BGC for each color of eggs. In order to determine the statistical significance of geographic variation in the level of difference of the mean BGC between blue and white eggs, we developed a permutation test, in which we first generated three random populations by shuffling the BGC, holding the observed sample size and egg color ratio of each population in our data set, and then we calculated the expected difference in the mean BGC between blue and white egg clutches in each population and finally the expected geographic variation in the BGC difference between two egg colors was determined. We repeated this procedure 10000 times, where statistical significance was determined by comparing the observed values to the 95% confidence interval of the expected values. We compared two of the three populations in each test, resulting in three different tests to compare all populations. All statistical analyses were conducted in R environments, version 2.14.1 (R Development Core Team, 2011).
Results
Spectrometry of egg color
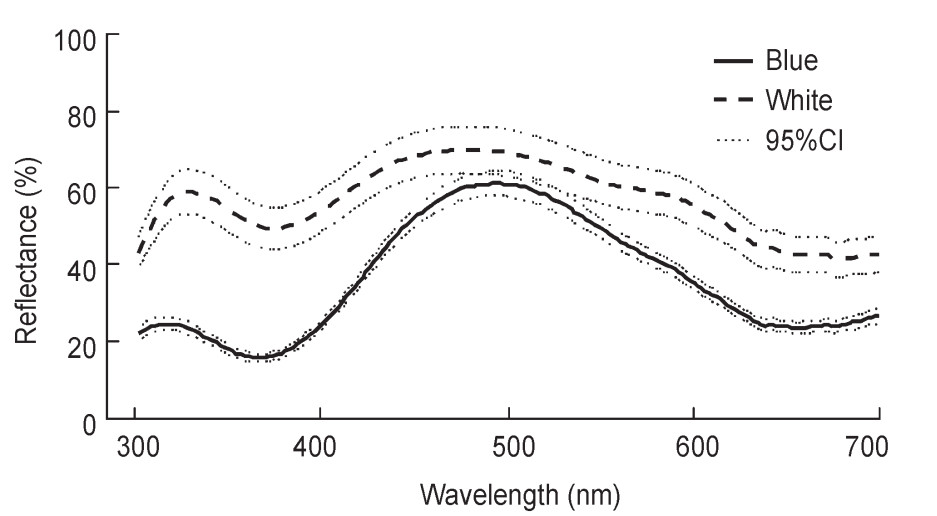
From measurements at three randomly selected spots for each egg, we found that the reflectance rate was highly repeatable not only within an egg but also within a clutch for both egg colors (within an egg, blue: r = 0.95, white: r = 0.94; within a clutch: blue: r = 0.78, white: r = 0.87), indicating consistent coloration within an egg as well as a clutch. Overall, the spectral shapes were consistent with the generally blue and white appearance to humans, although the spectral shapes of white eggs was slightly different from those expected from a pure white color (Fig. 2).
Figure
2.
Reflectance spectra of blue and white Vinous-throated Parrotbill eggs. Lines represent average reflectance spectra and the 95% confidence interval (CI), calculated from the average value of each clutch (blue= 38 clutches, white = 22 clutches).
Overall, the egg color ratios varied spatially among populations (Fisher's exact test, p < 0.001, Table 1). Interestingly, we found a geographic gradient in egg color ratios. That is, the frequency of nests with white eggs dramatically decreased as latitude declined towards the southern part of the Korean peninsula (from 58.6% at YM through 15.6% at GS to 4% at CM) (Table 1, Fig. 1).
Correlation of egg color variation and egg color ratio
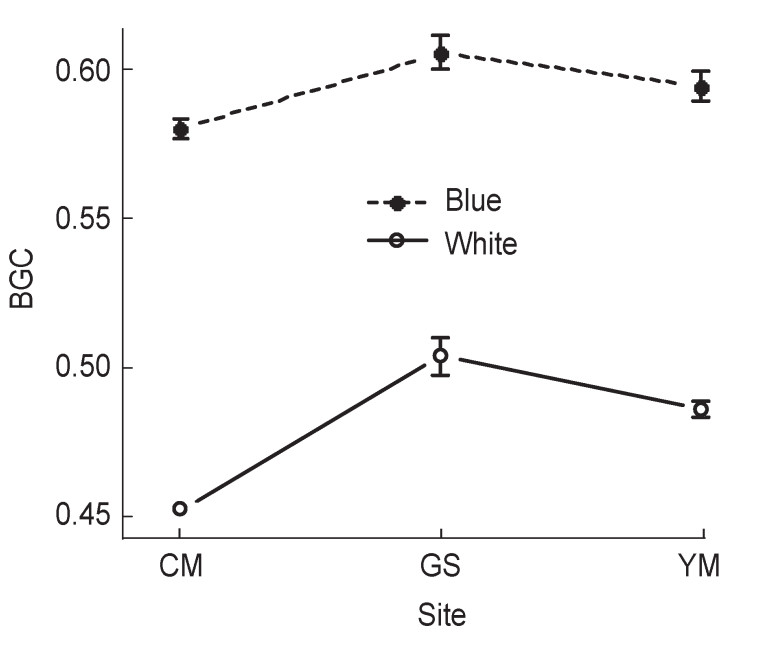
Blue and white eggs differed clearly in blue-green chroma (BGC) (Fig. 3). We found some geographical variation in BGC in both egg colors (blue egg: F = 7.76, df = 2, 34, p < 0.01; white egg: F = 8.62, df = 2, 19, p < 0.01, Fig. 3). However, these variations were not consistent with the variation in egg color ratios (average % blueegg clutch: 66.9% at YM, 84.4% at GS, 96% at CM). Furthermore, the level of difference in the mean BGC between blue and white eggs in each population did not differ among populations (permutation test, data not shown) (Fig. 3).
Figure
3.
Geographic variation in the intensity of blue-green chroma (BGC) of Vinous-throated Parrotbill eggs. Plots and whiskers indicate the mean and standard error of samples. Sample sizes of nests, where egg color was measured, are indicated in Table 1. Statistically significant comparisons are as follows: BGC- blue: YM vs. CM, p < 0.05; GS vs. CM, p < 0.001; white: YM vs. GS, p < 0.05; GS vs. CM, p < 0.001.
Monitoring egg color ratios over a large spatial-temporal scale would be fundamental to reveal the maintenance mechanism of egg color polymorphism (Liang et al., 2012). We found that overall egg color ratios were skewed considerably towards blue egg morphs, but the ratios were different geographically. Interestingly, we found a morph-ratio cline in egg color ratios; that is, the proportion of white eggs in a population dramatically decreased at lower latitudes. Morph-ratio clines have been observed frequently in birds with polymorphic appearances (Wunderle, 1981; Atkinson and Briskie, 2007) but no such cases have been reported with repect to egg colors. Our results are therefore the first showing the presence of a morph-ratio cline in polymorphic egg colors of birds. It is known that a selection gradient or gene flow, or an interaction of both, could generate such a morph-ratio cline in polymorphic species (Endler, 1977; Wunderle, 1981). Based on this knowledge, two potential scenarios could be proposed to account for the observed geographic gradient in egg color ratios.
First, the current egg color ratios may represent an equilibrium state at a certain range and reflect a selection gradient, such as geographic variation in brood parasitism and predation rate, the strength of which can vary over time and space. Cuckoo parasitism could be a major selective factor generating geographic variation in egg color ratios. In Ashy-throated Parrotbills, Yang et al. (2010) showed different egg color ratios from this study, with an even or weak bias towards white egg morphs. In that system, both Ashy-throated Parrotbills and Common Cuckoos have evolved the same pattern of egg color polymorphism, with hosts showing different rejection rates according to the mimetic levels of parasitic eggs. They proposed that the observed egg color ratio resulted from the frequency-dependent selection on egg colors between the host and parasite. In Korea, cuckoos using the Vinous-throated Parrotbill as a host lay blue eggs only and hosts with blue eggs reject cuckoo eggs with lower probability than those laying white eggs (Lee and Yoo, 2004). Unlike a system of Ashy-throated Parrotbills and Common Cuckoos in which the egg color ratio seems to be more or less at equilibrium by frequency-dependent selection, this situation would generate directional selection in the egg color ratio in a broad sense, so, for example as the parasitism rate increases in a population, the proportion of females laying white eggs could increase in the population. Therefore, our result may indicate that cuckoo parasitism may be more severe in the northern part of Korea than in the south if this scenario is correct.
Predation is also known as a primary selective pressure varying egg coloration in birds (Blanco and Bertellotti, 2002; Kilner, 2006) and therefore, if the relative predation rate according to egg colors is different geographically, it could lead to spatial variation in egg color ratios. In small passerines breeding with open-cup nests, however, nest predation rate appears not to be affected by egg colors as much as we believe (Götmark, 1992; Weidnger, 2001). Previous studies in this species also reported that the predation rate was not different between two color morphs (Kim et al., 1995). Therefore it seems unlikely that the current variation in egg color ratios may reflect geographic differences in the relative predation rate between the two color morphs.
Alternatively, the latitudinal cline in egg color ratios may not result from a selection gradient but occur by non-adaptive mechanisms such as gene flow, or under the effect of both. Thus, the proportion of white eggs in a population may represent the timing of the spread of the white color gene along the Korean peninsula. It may therefore be assumed that the white-egg gene has only recently reached the southern parts of the peninsula. Furthermore, this may indicate that the current egg color ratios could be viewed as a dynamic state with a potential to be changed rather than at an equilibrium state. It is generally admitted that gene flow within and among populations may be less restricted in birds than in other animal taxa due to their superior flight ability (Avise, 1996; van Treuren et al., 1999; Crochet, 2000). However, Lee et al. (2010) showed a considerable genetic differentiation among closely located winter flocks (< 5 km) in Vinous-throated Parrotbills and suggested that this limited gene flow, as a result of their philopatric nature, is likely to be the norm in this species. Consequently, if the gene for white color morph migrates down the Korean peninsula, this restricted gene flow may slow down the speed of gene spread, differentiating the existing period of white egg genes among populations. This, together with the directional selection in relation to cuckoo parasitism, may result in the clinal variation of egg color ratios along the Korean peninsula. Therefore, it seems unlikely that the degree of brood parasitism is the only factor determining egg color ratio of host populations. Instead, we may assume that both selection by cuckoos and gene flow generate the current morph-ratio cline in egg color ratios across the Korean peninsula. However, in order to clarify fully which scenario is correct, a long-term study monitoring egg color ratios and cline shift is needed.
Correlation of egg color variation and egg color ratio
Blue and white eggs are clearly different in their spectral shapes as well as in BGC. Each egg was immaculate and consistent in its color and intra-clutch variation in egg coloration was also very low in both blue and white egg clutches. We found some significant variation in BGC among the three populations, but the pattern of variation was not linked with the change in egg color ratios. In other words, we predicted that blue color should be bluer as the proportion of blue-egg nests decreases in a population, but this was not the case. Furthermore, we expected that the difference in BGC between blue and white eggs should be larger in a population with lower proportion of blue-egg nests but we did not find such differences among populations. Given that egg color ratios may reflect the degree of brood parasitism, this result seems to imply that brood parasitism by cuckoos may not be the major factor behind the variation in egg coloration among populations. Instead, the observed variation in BGC may simply be associated with local environmental conditions such as food availability and genetic compositions.
Potential scenario for the evolution of egg color polymorphism in Vinous-throated Parrotbills
According to Takasu(2003, 2005) and Yang et al. (2010), the evolution of egg color polymorphism by disruptive selection should be accomplished by a gradual shift in the system of avian brood parasitism, hence Yang et al. (2010) proposed that pale blue eggs in Ashythroated Parrotbills might be an original morph before splitting into two distinct colors: blue and white. A key prediction from this view is that the variation of egg color should be related to the strength and/or history of brood parasitism. However, we found no correlation between egg appearance and the degree of brood parasitism which was assumed by egg color ratios. Therefore, we cannot conclude whether disruptive selection is the only mechanism that generates the evolution of egg color polymorphism observed in the Korean population of the Vinous-throated Parrotbill.
Mutation and strong positive selection leading to fixation could be another way that a new trait evolves as shown in the example of Pepper Moths (Biston betularia) (van't Hof et al., 2011). In birds, the ancestral egg color was probably white (Kilner, 2006) and erratic occurrence of white mutant egg color seems not to be uncommon in nature (Gross, 1968). Thus, if this mutant white egg is beneficial with a large fitness reward for certain reasons, there may be a chance that it could be selected for and fixed in a population. Furthermore, the new trait could be spread among populations via gene flow. One possible scenario for the evolution of egg color polymorphism in the Vinous-throated Parrotbill is as follows; the mutant for white egg occurred in some places, which had considerable fitness benefits against cuckoo parasitism and thus survived in a population. Finally, the gene for white eggs has spread across the range of the species via a gene flow. As we discussed earlier, the clinal change in egg color ratios may imply that the gene producing white eggs in Korea did not evolve but immigrated from outside the Korean peninsula and is still spreading. Furthermore, the fact that white eggs are not or rarely recorded in some populations of Vinous-throated Parrotbills located at the boundary of the species range such as Taiwan, eastern Manchuria and southern Korea (Robson, 2007) may indicate that the gene has not reached into those areas yet. Then, where did the white mutant originate? Interestingly, in contrast to the Vinous-throated ParrotbillCommon Cuckoo system in Korea where only one host species lay polymorphic eggs, in the system of Ashythroated Parrotbills and Common Cuckoos in China, both interacting species lay polymorphic eggs in colors: blue, pale blue and white (Yang et al., 2010). This latter study was carried out in Guizhou province (28° 10'N, 107°10'E), China, which is near Sichuan province which is the center of distribution of the species in the family Paradoxornithidae (Robson, 2007). This result may indicate that the Ashy-throated ParrotbillCommon Cuckoo system in Guizhou province has advanced one step further than the system of the Vinousthroated Parrotbill and Common Cuckoo in Korea. In other words, this may imply that the parrotbill-cuckoo interaction has a longer history in Guizhou than in Korea. Furthermore, Yang et al. (2010) found an opposite egg color ratio to the one we found; that is, white egg clutches were more frequent than blue egg ones, implying that the presence of white egg clutches has a longer history in Guizhou province than in Korea.
Taken all these together, we can assume that the white egg morphs were generated by either disruptive selection or mutation somewhere in central China where the center of distribution of this species lies and that this new phenotype has spread its range with a greater fitness against cuckoo parasitism than that of the original blue eggs. Thus, we now may be observing an ongoing process of gene flow in the Korean peninsula. Although our hypothesis was formulated based on the results from some populations along the Korean peninsula, this area occupies only a small part of the range of the species. Furthermore, it is based on the assumption that the egg color ratio of a population reflects the degree of brood parasitism in the population but we cannot rule out the possibility that egg color ratios may not represent the degree of brood parasitism properly. Accordingly, our view is open to be altered until we have better data on brood parasitism rates and egg coloration. Further studies focusing on collecting egg color data across a larger geographical scale of the range of this species would be worthwhile in order to clarify further the evolutionary origin of egg color polymorphism in Paradoxornis species.
Acknowledgments
We are indebted to Sei-Woong Choi, YongNam Moon, Chang-Ku Kang, Myun-Sik Kim and Kyung-Gyu Lee for their valuable helps in the field and laboratorium. We also thank two anonymous reviewers, and the guest editors for helpful comments on our manuscript. This work was financially supported by Brain Korea 21 Fellowships.
Atkinson K, Briskie JV. 2007. Frequency distribution and environmental correlates of plumage polymorphism in the grey fantail Rhipidura fuliginosa. New Zeal J Zool, 34:273–281.
Avise JC. 1996. Three fundamental contributions of molecular genetics to avian ecology and evolution. Ibis, 138:16–25.
Blanco G, Bertellotti M. 2002. Differential predation by mammals and birds: Implications for egg-color polymorphism in a nomadic breeding seabird. Biol J Linn Soc, 75:137–146.
Collias EC. 1993. Inheritance of egg-color polymorphism in the village weaver (Ploceus cucullatus). Auk, 110:683–692.
Endler JA. 1977. Geographic Variation, Speciation, and Clines. Princeton University Press, Princeton, New Jersey.
Ford EB. 1945. Polymorphism. Biol Rev, 20:73–88.
Galeotti P, Rubolini D, Dunn PO, Fasola M. 2003. Color polymorphism in birds: causes and functions. J Evol Biol, 16:635–646.
Gigord LDB, Macnair MR, Smithson A. 2001. Negative frequency-dependent selection maintains a dramatic flower color polymorphism in the rewardless orchid Dactylorhiza sambucina (L. ) Soò. Proc Natl Acad Sci USA, 98:6253–6255.
Götmark F. 1992. Blue eggs do not reduce nest predation in the song thrush, Turdus philomelos. Behav Ecol Sociobiol, 30:245–252.
Gray SM, McKinnon JS. 2007. Linking color polymorphism maintenance and speciation. Trends Ecol Evol, 22:71–79.
Gross AO. 1968. Albinistic eggs (white eggs) of some North American birds. Bird Banding, 39:1–6.
Hoffman EA, Blouin MS. 2000. A review of color and pattern polymorphisms in anurans. Biol J Linn Soc, 70:633–665.
John RT. 2003. Insect melanism: The molecules matter. Trends Ecol Evol, 18:640–647.
Jones JS, Leith BH, Rawlings P. 1977. Polymorphism in Cepaea: A problem with too many solutions? Annu Rev Ecol Syst, 8:109–143.
Jonsson B, Jonsson N. 2001. Polymorphism and speciation in Arctic charr. J Fish Biol, 58:605–638.
Kilner RM. 2006. The evolution of egg color and patterning in birds. Biol Rev, 81:383–406.
Kim C-H, Yamagishi S, Won P-O. 1995. Egg-color dimorphism and breeding success in the crow tit (Paradoxornis webbiana). Auk, 112:831–839.
Lee J-W, Simeoni M, Burke T, Hatchwell BJ. 2010. The consequences of winter flock demography for genetic structure and inbreeding risk in vinous-throated parrotbills, Paradoxornis webbianus. Heredity, 104:472–481.
Lee J-W, Yoo J-C. 2004. Effect of host egg color dimorphism on interactions between the vinous-throated parrotbill (Paradoxornis webbianus) and common cuckoo (Cuculus canorus). Korean J Biol Sci, 8:77–80.
Lee Y. 2008. Egg discrimination by the vinous-throated parrotbill, a host of the common cuckoo that lays polychromatic eggs. M. Sc. thesis. University of Manitoba, Winnipeg, Canada.
Liang W, Yang C, Stokke BG, Antonov A, Fossøy F, Vikan JR, Moksnes A, Røskaft E, Shykoff JA, Møller AP, Takasu F. 2012. Modelling the maintenance of egg polymorphism in avian brood parasites and their hosts. J Evol Biol, 25:916–929.
Moreno J, Lobato E, Morales J, Tomas G, Martinez-de la Puente J, Sanz JJ, Mateo R, Soler JJ. 2006. Experimental evidence that egg color indicates female condition at laying in a songbird. Behav Ecol, 17:651–655.
Ortega C. 1998. Cowbirds and Other Brood Parasites. The University of Arizona Press, Tucson, AZ.
R Development Core Team. 2011. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. http://www.R-project.org/.
Robson C. 2007. Family Paradoxornithidae (parrotbills). In: del Hoyo J, Elliott A, Christie DA (eds) Handbook of the Birds of the World, Volume 12. Lynx Edicions, Barcelona.
Roulin A. 2004. The evolution, maintenance and adaptive function of genetic color polymorphism in birds. Biol Rev, 79:815–848.
Siefferman L, Navara KJ, Hill GE. 2006. Egg coloration is correlated with female condition in eastern bluebirds (Silia sialis). Behav Ecol Sociobiol, 59:651–656.
Spottiswoode CN, Stevens M. 2011. How to evade a coevolving brood parasite: egg discrimination versus egg variability as host defences. Proc R Soc B, 278:3566–3573.
Takasu F. 2003. Co-evolutionary dynamics of egg appearance in avian brood parasitism. Evol Ecol Res, 5:345–362.
Takasu F. 2005. A theoretical consideration on co-evolutionary interactions between avian brood parasites and their hosts. Ornithol Sci, 4:65–72.
van Treuren R, Bijlsma R, Tinbergen JM, Heg D, van de Zande L. 1999. Genetic analysis of the population structure of socially organized oystercatchers (Haematopus ostralegus) using microsatellites. Mol Ecol, 8:181–187.
van't Hof AE, Edmonds N, Dalíková M, Marec F, Saccheri IJ. 2011. Industrial melanism in British peppered moths has a singular and recent mutational origin. Science, 332:958–960.
Weidnger K. 2001. Does egg color affect predation rate on open passerine nests? Behav Ecol Sociobiol, 49:456–464.
Wunderle JM Jr. 1981. An analysis of a morph ratio cline in the bananaquit (Coereba flaveola) on Grenada, West Indies. Evolution, 35:333–344.
Yang C, Liang W, Cai Y, Shi S, Takasu F, Møller AP, Antonov A, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010. Coevolution in action: Disruptive selection on egg color in an avian brood parasite and its host. PLoS ONE, 5: e10816.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds