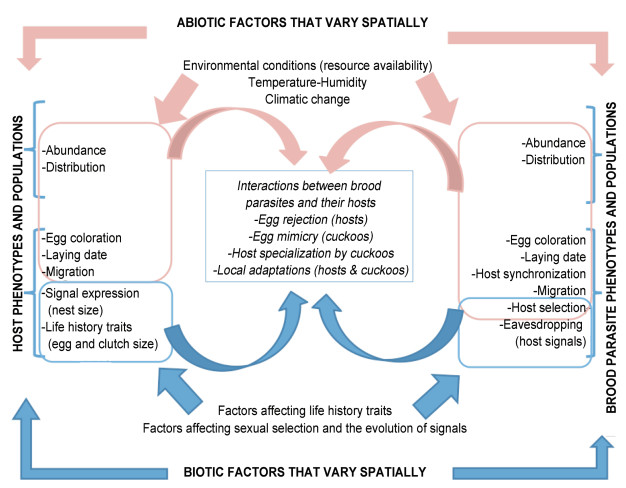
| Abiotic environment |
|
|
|
|
| Temporal pattern of mimicry in Reed Warbler |
No significant effect due to rainfall |
|
| Avilés et al.
(2006) |
| Temporal pattern of egg color in Reed Warbler |
A significant effect of rainfall on one component of host egg color |
| F = 6.23, df = 1, 458, p = 0.012 |
Avilés et al.
(2007) |
| Temporal pattern of egg color in Reed Warbler |
A significant effect of rainfall on one component of Cuckoo egg color |
| F = 5.21, df = 1, 457, p = 0.02 |
Avilés et al.
(2007) |
| Temporal pattern of mimicry in Reed Warbler and Cuckoo |
A significant effect of rainfall on one component of egg color matching |
| F = 4.81, df = 1, 459, p = 0.03 |
Avilés et al.
(2007) |
| Temporal pattern of mimicry in Reed Warbler |
A significant effect of rainfall on spottiness of egg color |
| F = 4.87, df = 1, 458, p = 0.03 |
Avilés et al.
(2007) |
| Temporal change in host use is linked to migratory phenology of Cuckoos and hosts |
A significant advancement in arrival date is linked to a climate warming |
| χ2 = 9.27, df = 1, p = 0.0023 |
Møller et al. (2011c) |
| Phenotype matching between eggs of Cuckoo and parasites among Reed Warbler populations |
Between population differences in host egg phenotype were related to differences in spring climatic conditions |
| Temperature: Mantel r = -0.64, p = 0.01; Rainfall: Mantel r = -0.87, p = 0.001 |
Avilés et al.
(2012) |
| Biotic environment |
|
|
|
|
| Nest size evolution due to sexual selection and brood parasitism |
Large nests signal phenotypic quality in magpies, and Great Spotted Cuckoos prefer hosts with large nests. However, in parasitized populations nest size has become small to prevent detection by cuckoos |
| Using parasitized nests: F = 24.87, df = 1, 308, p < 0.0001; Excluding parasitized nests: F = 19.67, df = 1, 268, p < 0.0001 |
Soler et al.
(1999a) |
| Life history of host and brood parasitism |
Increase in clutch size and decrease in egg volume in the presence of parasitism |
| F = 10.39, df = 1, 280, p = 0.0014; F = 10.53, df = 1, 262, p = 0.0013 |
Soler et al.
(2001a) |
| Life history of host and brood parasitism |
Clutch size was positively related to rejection rate, while egg size was negatively related to rejection rate |
| Mantel r = 0.33, p = 0.014; Mantel r = -0.38, p = 0.048 |
Soler et al.
(2001a) |
| Coevolutionary interactions |
| (a) Temporal patterns |
|
|
|
|
| Temporal pattern of parasitism |
Decrease in parasitism rate |
| χ2 = 47.1, df = 4, p < 0.001 |
Brooke et al.
(1998) |
| Temporal pattern of rejection |
Decrease in rejection rate |
| χ2 = 47.1, df = 4, p < 0.001 |
Brooke et al.
(1998) |
| Temporal pattern of rejection |
Increase in rejection from 13.8% to 89.3% in 16 years |
| χ2 = 49.15, df = 1, p < 0.0001 |
Robert and Sorci (1999) |
| Temporal pattern of parasitism rate |
Temporal increase in parasitism rate of Azure-winged Magpie |
| No test statistics reported |
Nakamura (1990) |
| Temporal pattern in mimicry |
Improved match with host eggs over time |
Match due to host choice |
F = 6.73, df = 1, 747, p = 0.018 |
Avilés et al.
(2006) |
| Temporal trend in parasitism and rejection rate |
Temporal increase in rejection rate over time |
| F = 6.07, df = 1, 31, p = 0.02 |
Soler et al.
(1994) |
| Temporal trend in parasitism rate |
Temporal increase in parasitized nests with cuckoo eggs laid by more than one female |
| F = 8.15, df = 2, 30, p = 0.017 |
Soler et al. (1994) |
| Temporal trend in parasitism rate |
Temporal increase in parasitized nests with more than one cuckoo egg |
| F = 4.22, df = 2, 123, p = 0.017 |
Soler et al. (1994) |
| Rate of parasitism and density of cuckoos |
Temporal increase in parasitized nests with increasing abundance of cuckoos |
| F = 18.16, df = 1, 31, p = 0.0002 |
Soler et al. (1994) |
| Ejection rate and abundance of cuckoos |
No significant increase in ejectio rate with abundance of cuckoos |
| F = 3.66, df = 1, 28, p = 0.07 |
Soler et al. (1994) |
| Ejection rate decreases in the absence of cuckoo parasitism |
Village Weaverbirds (Ploceus cucullatus) show increasing variance in introduced populations |
| Significant differences in between individual variation in Hispaniola (F = 3.83, df = 111, 143, p < 0.001, F = 5.81, df = 105, 153, p < 0.00001, F = 18.5, df = 3, 433, p < 0.001, F = 11.8, df = 111, 143, p < 0.001, F = 35.0, df = 3, 499, p < 0.001, F = 4.51, df = 105, 153, p < 0.00001, F = 7.5, df = 3, 431, p < 0.001, U = 8, 575, p < 0.001) and Mauritius (F = 3.26, df =106, 51, p = 0.00003, F = 18.5, df = 3, 433, p < 0.10, F = 3.26, df = 106, 51, p < 0.10, U = 3, 724, p < 0.10). More variable clutches in introduced populations in Hispaniola (F = 16.8, df = 3, 431, p < 0.0001, F = 19.9, df =143, 111, p < 0.001, F = 15.8, df = 3, 431, p < 0.001, F = 11.8, df = 143, 111, p < 0.001, U = 9.514, p < 0.001, U = 9, 665, p < 0.001, U = 8, 681, p < 0.001, U = 9, 657, p < 0.001) and Mauritius (F = 1.93, df = 51, 121, p < 0.05, F = 4.59, df = 51, 121, p < 0.05, U = 3, 624, p < 0.05, U = 3, 499, p < 0.10, U = 3, 532, p < 0.10) |
Lahti (2005) |
| Temporal pattern of parasitism rate by Brownheaded Cowbird |
Acadian Flycatcher (Empidonax virescens), Indigo Bunting (Passerina cyanea), Northern Cardinal (Cardinalis cardinalis) |
| Significant decline in parasitism rate and parasite intensity, although no test statistics reported |
Cox et al. (2012) |
| Parasitism rate |
Decrease in parasitism rate by Rufous-bush Robin |
| Kendall tau = −1.00, p = 0.008 |
Soler et al. (2012) |
| Ejection rate |
Decrease in rejection rate by Rufous-bush Robin |
| Kendall tau = −0.98, p = 0.001 |
Soler et al. (2012) |
| (b) Spatial patterns |
|
|
|
|
| Spatial pattern of rejection |
Higher level of rejection by village weavers in African range than in introduced range |
| No test statistic reported |
Cruz and Wiley (1989) |
| Spatial differences in parasitism and anti-parasite behavior |
Differences in egg rejection of cuckoo eggs among sites differ in relation to duration of sympatry |
| G = 31.47, df = 2, p < 0.001 |
Soler and Møller (1990), Soler (1990) |
| Spatial differences in parasitism and anti-parasite behavior |
Differences in egg rejection of Cowbird eggs among sites differ in relation to duration of sympatry |
Difference in rejection between two sites |
No test statistics reported |
Briskie et al. (1992) |
| Spatial differences in parasitism and anti-parasite behavior |
Differences in aggressive behavior towards Cowbird model in relation to duration of sympatry |
Difference in aggressive behavior by host between two sites |
No test statistics reported |
Briskie et al.(1992) |
| Spatial heterogeneity in parasitism rate |
Differences in parasitism rate and rejection rate |
Differences in rejection rates among three populations |
Azumino vs Nagano χ2 = 1.38, df = 1, p = 0.17); Azumino vs Noveyama (χ2 = 12.0, df = 1, p < 0.001); Nagano vs Noveyama (χ2 = 18.1, df = 1, p < 0.001) No test statistics reported for differences in parasitism rates |
Nakamura et al. (1998) |
| Spatial differences in parasitism |
Lower parasitism rates in small marginal populations of Reed Warblers |
| t = −3.46, df = 18, p < 0.005 |
Lindholm(1999) |
| Spatial differences in antiparasite behavior |
Lower rejection rates in small marginal populations of Reed Warblers |
| G = 18.9, df = 1, p < 0.0001, G = 2.5 df = 1, p = 0.11, G = 38.3, df = 1, p < 0.0001, G = 12.2, df = 1, p < 0.001, G = 0.2, df = 1, p = 0.6, G = 3.6, df = 1, p = 0.06, G = 0.2, df = 1, p = 0.6, G = 0.6, df = 1, p = 0.50 |
Lindholm(1999) |
| Spatial differences in antiparasite behavior |
Lower responses in populations with less parasitism |
| H = 25, 6, p < 0.0001, H = 15.3, p = 0.0008, G = 2.2, p = 0.33, G = 10.0, p = 0.0067, G = 0.4, p = 0.81, G = 0.9, p = 0.64 |
Lindholm(2000) |
| Spatial differences in parasitism and anti-parasite behavior |
Comparison of upland and marsh habitats |
| No test statistics reported |
Strausberger(2001) |
| Spatial heterogeneity in parasitism rate |
Differences in parasitism rate and rejection rate |
Difference in rejection between two populations |
Fisher's exact test, two tailed, p = 0.023; No test statistics reported for differences in parasitism rates |
Moskát et al.(2002) |
| Spatial differences in parasitism rate |
Differences in parasitism rate of Contopus flycatchers |
| No test statistics reported |
Underwood et al. (2004) |
| Frequency of parasitism |
Local spatial variation in parasitism rate related to density of Cowbirds |
| r = 0.79, p = 0.02 |
Jensen and Cully (2005) |
| Frequency of parasitism |
Cowbird parasitism increased with farm and house density |
| No test statistics reported |
Tewksbury et al. (2006) |
| Host density and cuckoo parasitism |
Parasitism only occurs in high density populations of hosts above a certain threshold |
| χ2 = 4.37, df = 1, p = 0.037 |
Stokke et al.(2007) |
| Spatial variation in cowbird parasitism |
Increase in rate of parasitism with abundance of Cowbirds |
| F = 97.29, df = 1, 14, p < 0.001 |
Jewell and Arcese (2008) |
| Spatial heterogeneity in parasitism rate |
Differences in parasitism rate and rejection rate |
No difference in rejection rates of three populations not associated with parasitism rates |
Fisher's exact test, two tailed, p > 0.53; No test statistics reported for differences in parasitism rates |
Moskát et al.(2008) |
| Spatial heterogeneity in distribution and host races |
Host races mainly occur in hosts with a homogeneous spatial distribution |
| t = 2.49, df = 63, p = 0.016 |
Soler et al.(2009) |
| Spatial patterns of population density and host races |
Host races mainly occur in hosts with a high density |
| t = 2.35, df = 63, p = 0.22 |
Soler et al.(2009) |
| Rejection rate and parasitism rate in Reed Warbler populations |
Increased rejection rate with increasing parasitism rate |
| Mantel r = 0.281, p = 0.007 |
Stokke et al.(2008) |
| Local adaptation of phenotype of cuckoo eggs to Reed Warbler host |
Cuckoo eggs did not match eggs of local hosts, and matching was lower in populations with more different host species |
| Local adaptation: F = 1.24, df = 1, 3260, p = 0.26
No. host species: Mantel r = 0.55, p = 0.013 |
Avilés et al.(2011) |
| Local adaptation of eggs of three hosts races of Cuckoos to three sympatric species of hosts |
Eggs of host races of Cuckoo matched those of the host in terms of size, spotting pattern and egg color |
| size: χ2 = 520.8, df = 2, p < 0.0001 spotting: F = 4.46, df = 2, 303, p = 0.012 chroma: F = 3.27, df = 2, 94, p = 0.04 |
Antonov et al.(2010) |
| Tolerance of Cuckoo parasitism and rates of parasitism among sites |
Tolerance increased with rate of parasitism |
| t = 4.65, df = 7, p = 0.0023 |
Soler et al.(2011) |
| Distribution of color pattern in the chaffinch becomes increasingly dissimilar to the color distribution of the Brambling towards the core area of breeding of the Cuckoo host race |
Spatial variation in distribution of egg color |
| Chaffinch: UK and Fennoscandia: latitude: F = 43.1, df = 1, p = 0.0001, longitude: F = 32.6, df =1, p = 0.0001, Fennoscandia: latitude: F = 16.5, df = 1, p = 0.0001, longitude: F = 8.31, df = 1, p = 0.0022, Sweden and Denmark: latitude: F = 17.1, df = 1, p = 0.0001, longitude: F = 10.8, df = 1, p = 0.0004. Brambling: latitude: F = 1.70, df = 1, p = 0.18, longitude: F = 0.42, df = 1, p = 0.59. Cuckoo: latitude: F = 1.03, df = 1, p = 0.33, longitude: F = 0.57, df = 1, p = 0.54) |
Vikan et al.(2011) |
| Phenotype matching between eggs of Cuckoo and parasites among Reed Warbler populations |
Between population differences in egg rejection were related to differences in egg matching |
| Mantel r = −0.73, p = 0.003, Mantel r = −0.62, p = 0.016, Mantel r = 0.80, p = 0.007 |
Avilés et al.(2012) |
| Phenotype matching between eggs of Cuckoo and parasites among Reed Warbler populations |
Between population differences in host egg phenotype were related to differences in parasitism rate |
| Mantel r = 0.56, p = 0.01 |
Avilés et al.(2012) |
| Differences in aggressive behavior among sites |
Aggressive behavior by hosts is more intense at sites with more Cuckoos |
| F = 10.5, df = 1, 30, p = 0.003, F = 9.2, df = 1, 30, p = 0.005, z = 2.35, p = 0.018, F = 10.2, df = 1, 29, p = 0.003 |
Langmore et al. (2012) |
| Differences in rejection rate and micro-satellite marker |
Differences in rejection rate of Cuckoo eggs by magpies depend on allele size in a micro-satellite marker |
| Mantel r = 0.88, p < 0.001 |
MartínGálvez et al.(2006) |
| Differences in rejection rate and parasitism rate |
Higher rejection rate in populations with higher parasitism rate |
| Mantel r = 0.62, p = 0.006 |
MartínGálvez et al.(2007) |
| Differences in allele size and parasitism rate |
Higher frequency of allele in populations with higher parasitism rate |
| Mantel r = 0.68, p = 0.002 |
MartínGálvez et al.(2007) |
| Rejection behavior and gene flow |
Rejection rate depends on gene flow from other populations of hosts |
| Mantel r = 0.64, p = 0.016 |
Soler et al.(1999b) |
| Parasitism and rejection of cuckoo eggs in relation to coevolutionary interactions within the focal study area and in neighboring study areas |
Positive relationship between rejection rate and parasitism among populations, even after controlling for gene flow among populations |
| Mantel r = 0.37, p = 0.011 |
Soler et al.(2001b) |

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds