Research Institute of Forest Ecology, Environment and Protection, Key Laboratory of Forest Protection of the State Forestry Administration, Chinese Academy of Forestry, Beijing 100091, China
2.
Northeast Institute of Geography and Agricultural Ecology, Chinese Academy of Sciences, Changchun 130012, China
3.
College of Wildlife Resources, Northeast Forestry University, Harbin 150040, China
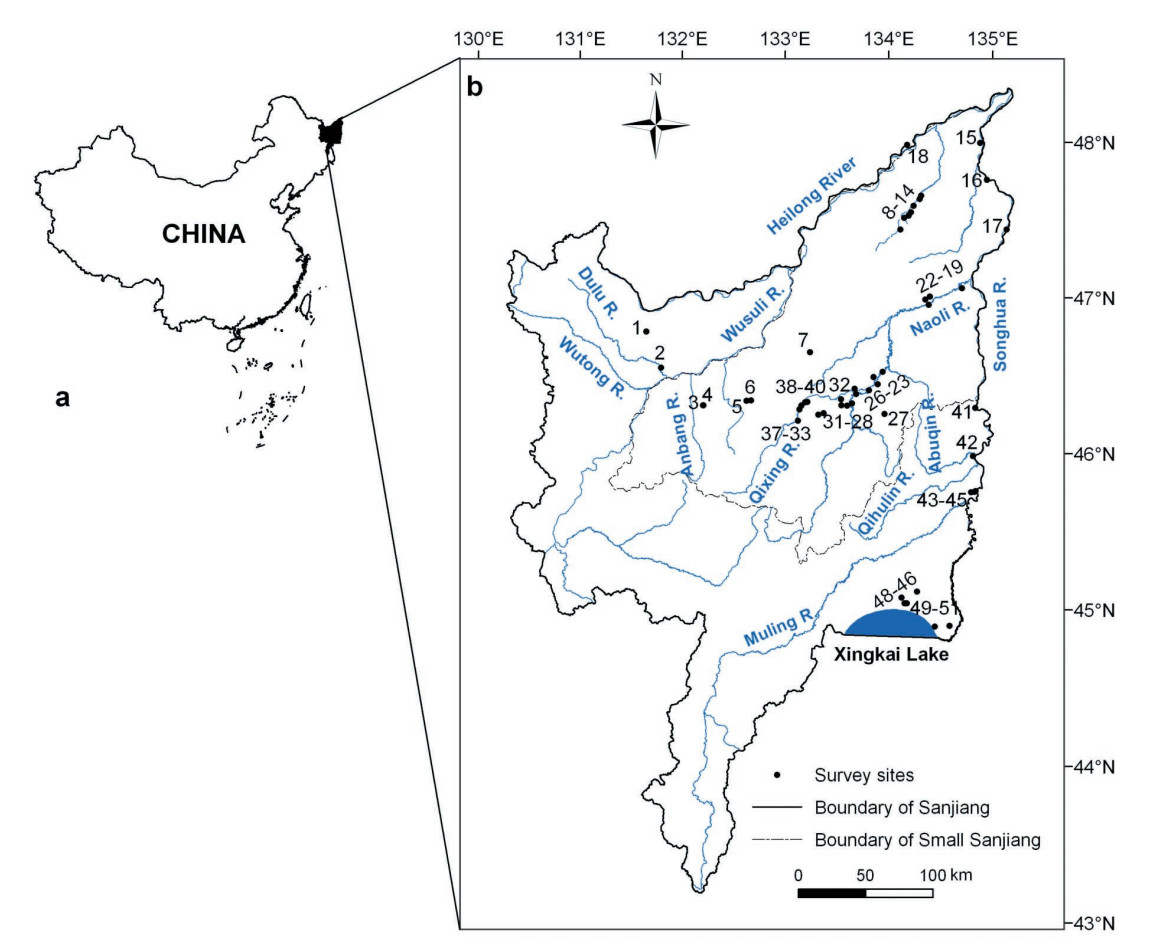
In order to measure the impact of changes in the marsh on breeding Red-crowned Cranes (Grus japonensis) and White-naped Cranes (G. vipio) in the Sanjiang Plain, we conducted a complete crane survey in each of the summers of 2007 and 2008, and obtained information on the changes in its marsh through interpretation of satellite images in 1986, 1995, 2000 and 2005. A comparison between the 1984 and 2008 census shows that: 1) the number of Red-crowned Cranes remained stable at about 300 birds, while the number of White-naped Cranes increased markedly; 2) the estimates of nests of Red-crowned cranes are close to the numbers of 1984, while approximately 90% of the nests of these two crane species were distributed over five national nature reserves (NNRs); 3) the two crane species became more concentrated in three regions, i.e., the Fuyuan Delta, the middle reaches of the Naoli and Qixing rivers, and the Xingkai Lake wetlands, rather as formerly in six regions. The area of the marsh decreased by 45% within the 20-year period from 1986 to 2005. The major loss of the marsh area was in the form of a conversion to paddy fields and dry land, which can be primarily attributed to direct human activity under various economic and policy drivers. The disappearance of cranes in the Dulu and Abuqin rivers was induced by large-scale agricultural development. The fragmentation into and isolation of small areas of the marsh became increasingly more serious, which resulted in unsuitable condtions for crane breeding in small patches of the marsh. The number of cranes declined from 1984-1995 but had recovered remarkably by 2008, which may be due to the establishment of a system of nature reserves, in place since 2000. The land ownership of the reserve plays a key role in maintaining the integrity of the wetland ecosystem, especially in Sanjiang, Honghe, Naolihe and Qixinghe NNRs, which together accommodated approximately 80% of the population of cranes. However, we did not discover any nests at all in the provincial and municipal reserves, which may be attributed to high human disturbances in these areas. In the end, we propose some recommendations on reserve management.
The Malayan Night Heron (Gorsachius melanolophus) inhabits a range of environments from India and Sri Lanka to Southeast Asia, southern China, Malaysia, Thailand, Taiwan, Indonesia, the Philippines and Japan (del Hoyo et al., 1992; The Ornithological Society of Japan, 2000). This species prefers tropical and subtropical wet forests as its breeding habitats (Hancock et al., 1978; del Hoyo et al., 1992). While it is a migratory bird in some areas (Hancock et al., 1978), it is a resident species in Japan (The Ornithological Society of Japan, 2000). Its distribution in Japan is usually restricted to the Yaeyama and Miyako islands, Okinawa Prefecture (Kawakami and Fujita, 2005). This heron is considered to be uncommon throughout much of its range (Kushlan and Hancock, 2005) and is classified as a vulnerable species in the Japanese Red List released in 2006. Understanding the diet of such a species is a very important issue for conservation because it relates closely to a species' ecological niche.
Little is known about the ecology of this heron species and only piecemeal information on its diet has been available, as this species breeds singularly, does not make a colony or communal roost, and inhabits shadowy forests. Shen and Chen (1996) described that 93% of the diet of the Malayan Night Heron were earthworms based on direct observation. Kushlan and Hancock (2005) estimated that the herons specialized in capturing frogs and very large earthworms, and also preyed on beetles, mollusks, crabs, snakes, lizards, birds and small fish. Regarding to the Japanese population, their diet has been seldom documented except for occasional reports (e.g. Makihara and Kawakami, 2005). However, no quantitative information of their food items has been available. The results of direct observations might bias towards large and noticeable objects at well-lighted area, and the proportion of inconspicuous items is very likely to be underestimated. To show a detailed dietary composition, we collected pellets regurgitated by the herons and stomach contents of dead individuals, because these analyses could provide quantitative estimation without the influence of ease of observation. In this paper, the dietary composition of the heron is described and its foraging microhabitats and circadian activity are discussed in light of dietary preference.
Methods
Pellets cast by Malayan Night Herons were collected under their roosts with the aid of mesh nets. Plastic mesh nets of 2 m × 2 m were located under the roost at a 1 m height from the ground and the pellets cast on the nets were collected after a period of days. The herons roost alone on trees at deciduous forests in the islands. The samples were kept in a freezer until the subsequent analysis. Collections were conducted in natural dense humid forests (Mt. Banna-dake) and riparian forests (Miyara River) on Ishigaki Island in March and April 2002 and in a secondary forest on Kohama Island in March 2003 (Fig. 1). The vegetation was mainly deciduous evergreen forest consisting of Castanopsis sieboldii, Quercus Miyagii, Hibiscus tiliaceus and Ficus trees. Additionally, the stomach contents of five dead birds were analyzed, four of them found on Ishigaki Island and one on Panari Island (Table 1). Three carcasses collected on Ishigaki Island were roadkill and one was rescued infirm. We extracted fragmented animal remains from the pellets and stomach contents, identified them to the lowest possible taxonomic level with the stereomicroscope, and counted the minimum number of each species found in each pellet.
Figure
1.
Map of the Yaeyama Islands, Japan. The black circles show the locations of pellet collection survey.
Thirty-eight pellets were collected on Ishigaki Island and five on Kohama Island, respectively. All pellets included a mass of sands and gravels, which were estimated to be swallowed with soil animals. The undigested remains of land snails, chilopods, arachnids, crabs, insects and reptiles were found among them (Table 2). Forty-three individuals of snails were found in 47% of the analyzed pellets and all of them were Acusta despecta. Two individuals of Chilopoda were found in one pellet on Ishigaki Island. Eight whip scorpions, nine spiders and unidentified arachnids were found in 30% of the pellets. The whip scorpions were certain to be Typopeltis crucifer, as no other species has been observed on the island. Twenty-two crabs were detected from 47% of the pellets, while all of the identified crabs were freshwater and 19 of them were of the family Potamidae. At least 69 individuals of 21 species of insects were detected from 65% of the pellets. Blattaria, Elateridae and Scarabaeidae were dominant and covered 55% of the total number of individuals. Nine reptiles were detected in 19% of all pellets and the dominant species was an agama lizard, Japalura polygonata ishigakiensis.
Table
2.
The species composition found in the pellets and stomach contents of Malayan Night Herons. Each number shows the number of individuals for each food item. The number in brackets is the percent occurrence of samples containing items for each taxon.
In the stomach contents of five carcasses, an arm of a frog Fejervarya sakishimaensis and several insects were detected (Table 2). The frog was estimated to be an adult and about 10 cm in its entire length. The dominant insects were cockroaches and grasshoppers, which were each detected in four of the five carcasses.
Discussion
Land snails, freshwater crabs and insects were dominantly detected in the diet of Malayan Night Herons. In insect species, cockroaches, click beetles and scarabs were the major prey organisms found in the pellets, whereas the proportions of cockroaches and grasshoppers detected were high in the stomach contents of carcasses. Most of the carcasses were collected in open areas, while pellets were found in forests. As grasshoppers frequently inhabit grasslands, the differences among them were consistent with their habitat uses.
The land snails found in the pellets were major terrestrial species found on the forest floor of Ishigaki Island. Freshwater crabs inhabit streams and the ground nearby. The main microhabitats of the detected cockroaches, click beetles, whip scorpions, spiders, and Scolopendra were litter, soil, and fallen and decaying trees, indicating that the herons prefer to forage on soil animals on the forest floor. Though the agama lizard is arboreal, they often use ground for movement. The scarabs are arboreal and aerial, but observing them walking along the ground is not uncommon. Since the night herons were unlikely to forage on trees, they probably occasionally preyed on scarabs on the ground. Such arboreal animals are suggested to contribute the diet menu of the birds as well as soil animals. Especially, the contribution of the lizards ought to be larger than the other species due to their larger body masses.
Acusta despecta detected in the pellets has nocturnal activity as with other various terrestrial mollusks (Kurozumi, 1997; Cook, 2001). Freshwater crabs are primarily nocturnal and remain hidden during the day in sheltered places (Yeo et al., 2008). The identified Coleoptera, Orthoptera and Blattaria are also nocturnal, which get active from evening. On the other hand, the agama lizards are diurnal (Tanaka, 1986). The herons had been considered to be generally crepuscular or nocturnal, while diurnal activities have been frequently observed (Brazil, 1991; del Hoyo et al., 1992). The current results implied that the herons spend both day and night times for foraging in response to the prey activities.
Although Kushlan and Hancock (2005) suggested that the herons foraged on fish in aquatic environments, distinct aquatic animals were not detected in the current study, while freshwater crabs and partial aquatic species were frequently found. The beak of this heron is shorter and thicker than that of typical herons, while other heron species generally have straight and pointed bills (del Hoyo et al., 1992), which are considered to be adapted to preying on fish. The bill morphology of the Malayan Night Heron is similar to that of the Japanese Night Heron (Gorsachius goisagi), which is considered to be adapted for foraging on soil animals by digging (Kawakami et al., 2005). Freshwater crabs are frequently found in litter layers around streams. Chang (2000) also noted that the Malayan Night Heron rarely foraged on fish. The current result and the morphological character imply that the dominant diet of the Malayan Night Heron consists of small animals in litter layers.
On the other hand, some distinct items, such as aquatic animals, might have been overestimated in their diet menu in the previous studies. The main component of their pellets was sand and gravels, which made the pellets breakable and indistinctive. When the pellets would go into the ground, they ought to be easy to break into pieces. It becomes hard to detect the inconspicuous components such as soil insects due to their naturalness on the ground. Conversely, remains of aquatic animals should be distinct, because it is unnatural that such items are there. In order to avoid overestimation, pellet traps with plastic mesh nets are effective.
The herons appear to prefer wet ground, sometimes near streams, as foraging habitat, where they can capture land snails and freshwater crabs. Since we could not detect fully aquatic animals, the heron was suggested not to directly utilize aquatic habitats. They are believed to prefer moist forests associated with watery environments such as streams and swamps because of the availability of soil animals. These results are consistent with the foraging preferences of the congeneric Japanese Night Heron, which breeds on the mainland of Japan (Kawakami et al., 2005). To conserve the Malayan Night Heron, forests with well developed litter layers and fallen and decaying trees, which are generally rich in the biomass of soil animals, must be maintained.
Malayan Night Herons were described to mainly forage on earthworms based on direct observation (Shen and Chen, 1996; Chang, 2000), although we did not detect any because they have few indigestible parts. However, earthworms were also not observed in the carcasses analyzed in this study. Though the night herons should presumably forage on earthworms, the frequency is likely to vary according to habitat. Confirmation is needed as to how important earthworms are in the diet of the Malayan Night Heron. Combination of multiple methods is required in order to show the actual diet composition.
Frogs were not frequently detected in the current study, although Kushlan and Hancock (2005) estimated that they were one of the heron's main diet sources in forests. This might be caused by the above-mentioned overestimation of eye-catching items, while it is possibly attributed to seasonal variation of food availability. Herons generally have variable diets related to availability of prey in their habitat (del Hoyo et al., 1992) and can seasonally change their diet menus (e.g., Marquiss and Leitch, 1990; Rodriguez et al., 2007). Frogs become active during and after rain events in general and the biomass seasonally undergoes a complete change. Thus, although the herons may often forage on frogs in rainy seasons, all of the pellets analyzed in the current study were collected during the same period, i.e., spring. In addition to the seasonal variation of diet menu, the potential for variation exists depending on location. The dietary menu of the Malayan Night Heron in other seasons and places should be systematically investigated as the next step.
Acknowledgments
We are especially grateful to Taiji Kurozumi for identifying the snails, Tetsuya Watanabe and Yasuhisa Henmi for identifying crabs, Yuichi Ohba and Hitoo Ohira for identifying elaterids, and Akio Tanigawa for identifying arachnids, and for providing corresponding ecological information. We also thank Takashi Fujii, Takashi Hiraoka and the Yamashina Institute for Ornithology for providing the carcass samples, Ako Sukegawa, Minako Murakami, Nanami Kawamura, Ryoko Kanzawa, Yoko Hashiguchi and Yoichiro Sakiyama for collecting the pellets in the field, and Miyuki Mashiko for analyzing the samples.
BirdLife International. 2012. Grus japonensis and G. vipio. In: IUCN 2012. IUCN Red List of Threatened Species. Version 2012.1. . Accessed on 09 July 2012.
Bradter U, Gombobaatar S, Uuganbayar C, Grazia TE, Exo KM. 2005. Reproductive performance and nest-site selection of White-naped Cranes Grus vipio in the Ulz river valley, north-eastern Mongolia. Bird Conserv Int, 15: 313-326.
Cheng L, Li XM. 2006. The status and conservation of Cranes in Heilongjiang Province. Territ Nat Resour Study, 1: 90-91. (in Chinese with English abstract)
Feng KM, Li JL. 1985. Aerial surveys of the Red-crowned Cranes and other waterflowls in China. J Northeast Forest Univ, 13(1): 80-87. (in Chinese with English abstract)
Feng SZ, Liu HJ, Yu WT, Jiang ZD. 2005. The survey on bird diversity in Xingkai Lake, China. Wetland Sci, 3(2): 149-153. (in Chinese with English abstract)
Frederick PC, Towles T, Sawicki RJ, Bancroft TG. 1996. Comparison of aerial and ground techniques for discovery and census of wading bird (Ciconiiformes) nesting colonies. Condor, 98: 837-841.
Fujita G, Harris J, Bold A, Tseveenmayadag N, Chuluunbatar S. 1994. Habitat preference of Demoiselle and White-napped Cranes, Anthropoides virgo and Grus vipio, breeding in Mongolia. In: Higuchi H, Minton J (eds) The Future of Cranes and Wetlands. Wild Bird Society of Japan, Tokyo, Japan, pp 93-96.
Han YL, Ge DN, Zhang YX. 2003. The research about birds resource and conservation in Sanjiang Plain. Territ Nat Resour Study, 1: 76. (in Chinese with English abstract)
Hanski I. 1998. Metapopulation dynamics. Nature, 396: 41-49.
Harris J. 1994. Cranes, people and nature: preserving the balance. In: Higuchi H, Minton J (eds) The Future of Cranes and Wetlands. Wild Bird Society of Japan, Tokyo, Japan, pp 1-14.
Hilton-Taylor C. 2000. IUCN Red List of Threatened Species. Gland, Switzerland and Cambridge, UK: IUCN.
Hua Y, Li XM, Liu XC, Sun ZY. 2006. Comparison of bird diversity during spring and autumn in Honghe Nature Reserve. J Northeast Forest Univ, 34(3): 23-28. (in Chinese with English abstract)
Jiang HX, Liu CY, Qian FW, Li CY, Qiu FC. 2009. A model of nest-site selection of Red-crowned crane based on RS, GIS and GPS techniques at Zhalong wetland, China. Sci Silv Sin, 45(7): 76-83. (in Chinese with English abstract)
Lee SD, Jablonski PD, Higuchi H. 2007. Winter foraging of threatened cranes in the Demilitarized Zone of Korea: behavioral evidence for the conservation importance of unplowed rice fields. Biol Conserv, 139(1-2): 286-289.
Li FM, Li PX. 1998. A comparative study on territories of White-napped Crane and Red-crowned Crane. Acta Zool Sin, 44(1): 109-111. (in Chinese with English abstract)
Li FM, Li PX. 1999. A comparative study on territories of White-napped Crane and Red-crowned Crane. China J Ecol, 18(6): 33-37. (in Chinese with English abstract)
Li XM, Hu YM, Ma YJ, Jia JL. 2003. Status and conservation of cranes in Sanjiang Plain. Territ Nat Resour Study, 1: 74-75. (in Chinese with English abstract)
Liu HY, Li ZF. 2008. Effects of landscape change of wetlands on habitats of waterfowls within Honghe Nature Reserve by its surrounding area. Acta Ecol Sin, 28(10): 5011-5019. (in Chinese with English abstract)
Liu HY, Zhang SK, Li ZF, Lv XG, Yang Q. 2004. Impacts on wetlands of large-scale land-use changes by agricultural development: The small Sanjiang Plain, China. Ambio, 33(6): 284-288. (in Chinese with English abstract)
Liu XT. 1997. Marsh resource and its sustainable utility in the Songnen-Sanjiang Plain. Sci Geogr Sin, 16(Suppl. ): 451-460. (in Chinese with English abstract)
Liu XT. 2000. Influence of large-scale reclamation on natural environment and regional environmental protection in the Sanjiang Plain. Sci Geogr Sin, 20: 14-19. (in Chinese with English abstract)
Liu XT, Ma XH. 2002. Natural Environmental Changes and Ecological Protection in the Sanjiang Plain. Science Press, Beijing. (in Chinese)
Ma YQ, Jin LR. 1987. The numerical distribution of the Red-crowned Crane in Sanjiang Plain area of Heilongjiang Province. Acta Zool Sin, 33(1): 82-87. (in Chinese with English abstract)
Masatomi H, Ogawa S, Miyama K. 1990. The relations between land cover monitored by satellite remote sensing and distribution of the Tancho's nests in Kushiro Marsh, Hokkaido. J Hokkaido Coll, Sensu Univ, 23: 209-223.
Mcgarigal K, Cushman SA, Neel MC, Ene E. 2002. FRAGSTATS v3: Spatial Pattern Analysis Programme for Categorical Maps. Computer software programme produced by the authors at the University of Massachusetts, Amherst. . Accessed on 10 October 2008.
Noss RF, Murphy DD. 1995. Endangered species left homeless in sweat home. Conserv Biol, 9: 229-231.
Pang SL. 2000. The report on the aerial survey of Red-crowned Crane in the Zhalong Nature Reserve. J Sci Teach Coll Univ, 20(2): 59-60. (in Chinese)
Piao RZ, Han AH, Zhang MH. 2000. Remote sensing and geographic information system to estimate habitat selection of red-crowned crane in Sanjiang Plain of Northeastern China. In: China Ornithology Association, Wild Bird Society of Taipei and China Wildlife Conservation Association (eds) Studies on Chinese Ornithology — Proceedings of the 4th Ornithological Symposium of Mainland and Taiwan, China. China Forestry Publishing House, Beijing, pp 170-197. (in Chinese with English abstract)
Prugh LR, Hodges KE, Sinclair ARE, Brashares JS. 2008. Effect of habitat area and isolation on fragmented animal populations. PNAS, 105(52): 20770-20775.
Rodgers JA, Kubilis PS, Nesbitt SA. 2005. Accuracy of aerial surveys of waterbird colonies. Waterbirds, 28(2): 230-237.
Schmidt BD, Roland J. 2006. Moth diversity in a fragmented habitat: importance of functional groups and landscape scale in the boreal forest. Ecol Popul Biol, 99: 1110-1120.
Smirenski SM. 1988. Chick relationships and brood sizes in Red-crowned (Grus japonensis) and White-napped (Grus vipio) Cranes. In: Litvinenko NM, Neufeldt IA (eds). The Palearctic Cranes. Amur-Ussuri Branch of the USSE Ornithological Society, Vladivostok. USSR, pp 49-53.
Smirenski SM, Harris J, Gray C, Strom K, Ichida N. 1995. Report of the Amur program of the Socio-Ecological Union. In: Halvorson CH, Harris JT, Smirenski SM (eds) Cranes and storks of the Amur River: the Proceedings of the International Workshop. Arts Literature Publishers, Moscow, Russia, pp 205-212.
Song KS, Liu DW, Wang ZM, Zhang B, Jin C, Li F, Liu HJ. 2008. Land use change in Sanjiang Plain and its driving forces analysis since 1954. Acta Geogr Sin, 63(1): 94-104. (in Chinese with English abstract)
Su LY, Xu J, Zhou DS. 1991. Breeding habitats of White-napped Cranes at Zhalong Nature Reserve. In: Harris J (eds) Prodeedings of 1987 International Crane Workshop. International Crane Foundation, Baraboo, Wisconsin, USA, pp 51-57. (in Chinese)
Su LY, Wang QS. 2010. The mainland population of the red-crowned crane faces growing threat. China Crane News, 14(1): 3-8.
Swift TL, Hannon SJ. 2010. Critical thresholds associated with habitat loss: a review of the concepts, evidence, and applications. Biol Rev, 85: 35-53.
Temple SA. 1986. The problem of avian extinctions. Curr Ornithol, 3: 453-485.
Wang QS. 2008. Threats for Red-crowned Crane. China Crane News, 12(2): 7-12.
Yang XJ, Zhang SM, Zhang XX, Feng WY. 2005. The composing and analysis of large waterbirds diversity in the Sanjiang Plain. J Northeast For Univ, 33(3): 56-58. (in Chinese with English abstract)
Zhang GC, Deng W, Lu XG, Song KS, Li HD, Zhang HY. 2007. The dynamic change of wetland landscape patterns in Xinkai River Basin. J Nat Resour, 22(2): 204-210. (in Chinese with English abstract)
Zhang SQ, Na XD, Kong B, Wang ZM, Jiang HX, Yu H, Zhao ZC, Li XF, Liu CY, Patricia D. 2009. Identifying wetland change in China's Sanjiang Plain Using Remote Sensing. Wetlands, 29(1): 302-313.
Zhao KY (ed). 1999. Chinese Mire Records. Science Press, Beijing. (in Chinese)
Zhou DS, Su LY. 1990. Preliminary study on the territory behavior of Red-crowned Crane. In: Heilongjiang Forestry Department (ed) Conservation and Research of International Cranes. China Forestry Publishing House, Beijing, pp 54-58. (in Chinese)
Zou HF, Wu QM, Ma JZ. 2003. The nest-site selection of Red-crowned crane in Zhalong Nature Reserve after burning and irrigating. J Northeast Norm Univ, 35(1): 54-59. (in Chinese with English abstract)
Table
2.
The species composition found in the pellets and stomach contents of Malayan Night Herons. Each number shows the number of individuals for each food item. The number in brackets is the percent occurrence of samples containing items for each taxon.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds