
Figure
1.
A female (left) and male (right) Brown-headed
Cowbird (photo by J. Rivers)
| Citation: |
Iftikhar HUSSAIN, Asmat-un-NISA, Sangum KHALIL. 2012: Population biology of the Grey Francolin (Francolinus pondicerianus) in an agro-ecosystem of the Pothwar Plateau, Pakistan. Avian Research, 3(2): 91-102. DOI: 10.5122/cbirds.2012.0009

|
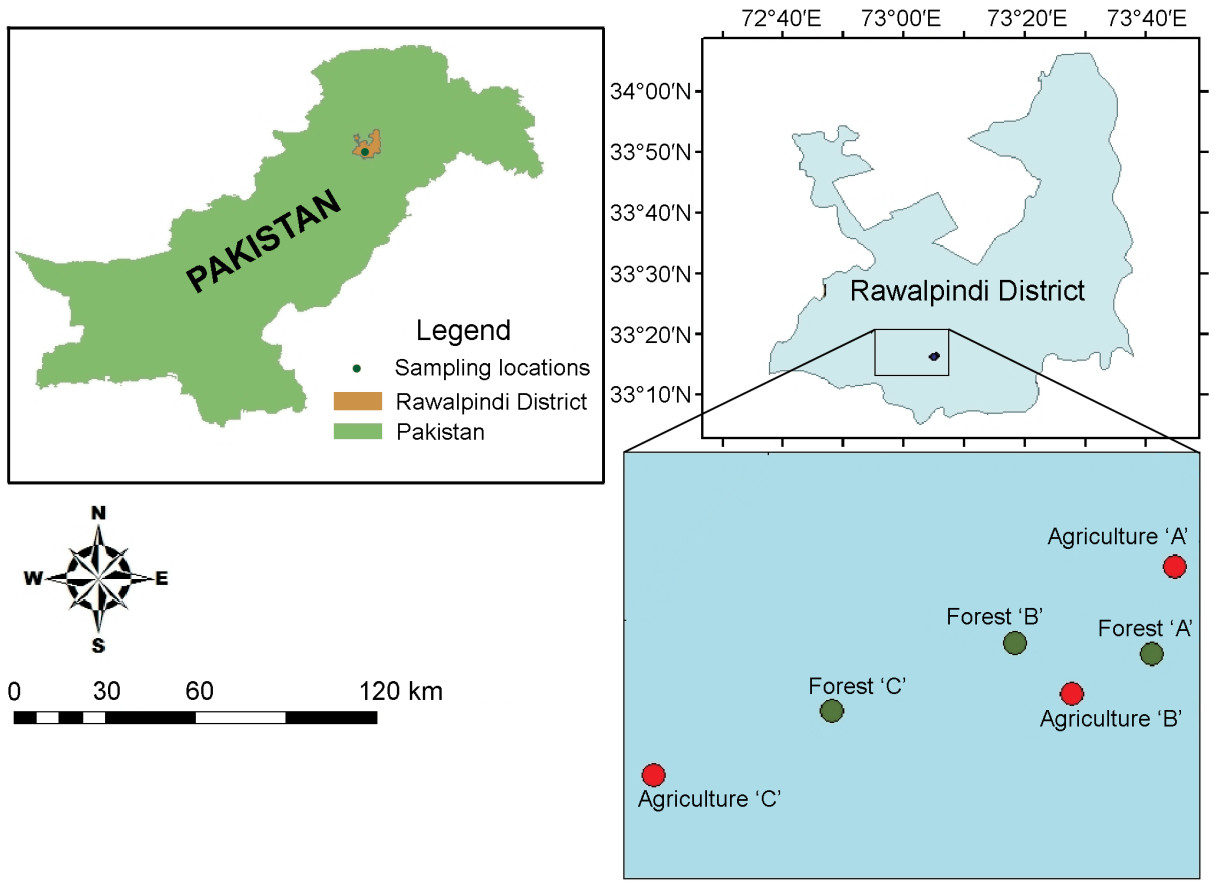
The field biology of the Grey Francolin (Francolinus pondicerianus) was studied in 2009 in an agro-ecosystem of the Pothwar area, an important range of its distribution in Pakistan. The bird density was estimated as 1.59±0.39 (mean±standard error) birds per ha in crop fields and 0.87±0.14 birds per ha in forest tracks, with some seasonal fluctuations in its habitat. All bird nests were located on the ground in vegetation mainly comprised of Desmostachia bipinnata, Acacia modesta, Imperata cylindrical, Zizipus jujuba and Euphorbia spp. Egg laying was recorded during the spring and summer months, with a mean egg laying span of 6±0.36 days (range 5-7 days) and a mean clutch size of 7±0.36 (range of 6-8 eggs). The mean incubation period was 20.6±0.50 days (range 19-22 days). Out of a total number of 42 eggs recorded, 32 hatched (76.19% success) with a mean hatching rate of 5.33±1.22 eggs per clutch. The fledgling success was estimated as 3.83±0.83 per clutch (63.08%). The species was found to be omnivorous and euryphagus in its feeding habit. There were 10 identifiable food items in the diet of the species, consisting of seven plant species (Pennisetum typhoideum, Sorghum bicolor, S. halepense, Prosopis juliflora, Phaseolus radiates, Carthemus axycantha and Acacia sp.), two insect species (Coptotermes formosanus and Formica rufa) and one grit.
The Brown-headed Cowbird (Molothrus ater; Figs. 1 and 2) is the only widespread brood parasite in the United States and Canada. The Bronzed Cowbird (M. aeneus) is restricted to the southern United States (Peer and Sealy, 1999a; Ellison and Lowther, 2009) and the Shiny Cowbird (M. bonariensis) is found in limited numbers in the southeastern United States (Reetz et al., 2010; Post and Sykes, 2011). It was believed that the Shiny Cowbird would rapidly invade and spread into the southeastern states (Cruz et al., 1998; 2000), but their range expansion has stalled for reasons that are unclear. The Bronzed and Shiny Cowbirds are found primarily in the Neotropics and this region also has the remaining two species in the parasitic cowbird clade, the Giant (M. oryzivorus) and Screaming (M. rufoaxillaris) Cowbirds. The latter is one of the most special-ized brood parasites in the world, primarily parasitizing a single species and on occasion four other species (DiGiacomo et al., 2010). In contrast, the Shiny Cowbird (266 hosts), followed closely by the Brown-headed Cowbird (247 hosts; Lowther, 2012), has the largest number of known host species of any brood parasite.

Cowbirds are considered grassland and edge species, but they also search for host nests in forested habitats. Brown-headed Cowbirds (hereafter cowbirds) commute large distances (7–15 km) on a daily basis between breeding and foraging locations (Dufty, 1982; Rothstein et al., 1984; Thompson, 1994; Curson et al., 2000). Their foraging habitat includes grasslands and pastures where they feed on seeds and invertebrates (Ortega, 1998), and they show a distinct preference for foraging in association with livestock which flush insects in these areas (Ortega, 1998). Historically, cowbirds followed and foraged in association with bison (Bison bison) herds (Mayfield, 1965). Their preference of open habitats has also impacted the evolution of host defenses, because all hosts known to eject cowbird eggs mostly nest in grasslands or open areas along edges apparently due to the fact these hosts have had the longest period of time to evolve defenses (Peer and Sealy, 2004).
The cowbird is a host generalist and usually parasitizes open-cup nesting species and avoids those that nest in cavities (Ortega, 1998; but see Peer et al., 2006; Hoover and Robinson, 2007). Cowbirds use hosts that feed their young insects (Ortega, 1998), but they also parasitize those with diets of seed and fruit despite the fact their young do not survive on such diets (Rothstein, 1976; Middleton, 1991; Kozlovic et al., 1996). Parasitism frequencies vary regionally, within hosts, and seasonally. For example in the fragmented forests of Illinois, parasitism frequencies are often 100% (Robinson, 1992; Fig. 3). Birds nesting at forest edges and in shrublands are parasitized at intermediate frequencies and those nesting in grasslands in the same region are rarely parasitized (Peer et al., 2000). In contrast, in the center of the cowbird's historic range in Kansas where the landscape consists almost wholly of grasslands, grassland species are parasitized at high frequencies (Elliott, 1978; Rivers et al., 2010). Parasitism of some host species, notably the Red-winged Blackbird (Agelaius phoeniceus) and Dickcissel (Spiza americana), also vary geographically (Linz and Bolin, 1982; Jensen and Cully, 1995; Searcy and Yasukawa, 1995) possibly due to alternative host choices within given avian communities (e.g., Barber and Martin, 1997).

Individual female cowbirds sometimes specialize on a particular host species and in other cases use multiple hosts (Alderson et al., 1999; Strausberger and Ashley, 2005), and host usage by individual female cowbirds is an area in need of further research. Estimates of the numbers of eggs laid by individual females in the wild range from 13 to more than 40 (Scott and Ankney, 1983; Fleischer et al., 1987; Alderson et al., 1999; see also Holford and Roby, 1993), but there is some consensus that cowbirds lay eggs on about 70–80% of the days during their breeding season (Fleischer et al., 1987). Multiple parasitism is relatively common in cowbirds (Robinson, 1992; Rivers et al., 2010; Fig. 4) and can result from multiple females parasitizing a nest or indi-vidual females laying multiple eggs within a single nest (Alderson et al., 1999; McLaren et al., 2003; Rivers et al., 2012).
In comparison to other brood parasite-host systems, cowbirds and their hosts have interacted for a relatively short time; the Molothrus clade is only 2.8–3.8 million years old (Rothstein et al., 2002). Perhaps as a result, the Brown-headed Cowbird lacks some adaptations possessed by other brood parasites (Mermoz and Ornelas, 2004) including mimicry of host eggs (Rothstein and Robinson, 1998; but see Peer et al. 2002), or evidence of specialized nestling adaptations such as directly killing nestmates (Lichtenstein and Sealy, 1998; Peer et al., 2013). Among the adaptations possessed by cowbird are that females have a larger hippocampus to remember where host nests are located (Sherry et al., 2003); laying eggs rapidly and before sunrise to avoid detection by hosts (Scott, 1991; Sealy et al., 1995; see also Peer and Sealy, 1999b); thick eggshells presumably to withstand puncture-ejection by hosts (Picman, 1989); greater pore diameter in their eggshells for increased embryonic respiration and shorter incubation periods (Jaeckle et al., 2012; see also Briskie and Sealy, 1990); removal of host eggs by females in conjunction with parasitism to enhance incubation efficiency (Peer and Bollinger, 1997; 2000) and for nutrition (Sealy, 1992); egg puncture and killing nestlings to force hosts to renest providing additional chances for parasitism (Arcese et al., 1996; Elliott, 1999; Hoover and Robinson, 2007; Dubina and Peer, 2013); and possibly forcing hosts to accept parasitism through mafia enforcement tactics (Hoover and Robinson, 2007).
Likewise, host defenses against cowbird parasitism appear to be relatively unsophisticated. For example, many hosts aggressively defend their nests against cowbirds, but the success of this strategy in preventing parasitism is limited (Sealy et al., 1998). Only approximately 10% of hosts reject cowbird eggs (Peer and Sealy, 2004; Fig. 5) and there is a bimodal response in that most hosts either accept or reject 100% of the time (Rothstein, 1975; Peer and Sealy, 2004), although more intermediate rejecters have been recently discovered (Peer et al., 2000, 2002, 2006). Smaller hosts are more likely to desert parasitized nests likely due to bill-size constraints that prevent egg ejection or simply because nest desertion evolves more easily than egg ejection (Hosoi and Rothstein, 2000). One host, the Yellow Warbler (Dendroica petchia), is unique because it often buries cowbird eggs in a new nest lining (Sealy, 1995) and it also has a specific referential call for cowbirds (Gill and Sealy, 2004). The lack of adaptive response to parasitism in newly parasitized hosts and some others appears to be due to an evolutionary lag (Rothstein, 1975; Peer and Sealy, 2004). Hosts with small bills that have difficulty in removing the cowbird egg or those with eggs that resemble cowbird eggs and may mistake them for their own may be in an evolutionary equilibrium (e.g., Rohwer and Spaw, 1988), but there is no compelling evidence of equilibrium among cowbird hosts to date (Peer and Sealy, 2004).
The Brown-headed Cowbird, and to a lesser extent the Shiny Cowbird, is unique among brood parasites because there are control programs designed to aid endangered species by culling cowbirds. To some extent, these control measures have been implemented based on misconceptions concerning impacts cowbirds have at the level of host populations and on incorrectly assuming that cowbirds rather than anthropogenic habitat destruction are limiting host populations (Rothstein and Peer, 2005). Unfortunately, there is no indication that control measures will cease despite some host populations having surpassed mandated minimum population goals and the expenditure of scarce management funds that could be put to better use (Hammer, 2011, unpubl. report). The management debate is exacerbated by the negative attitudes directed towards cowbirds by both birdwatchers and scientists (Ortega, 1998; see Peer et al., 2013, pers. observ.) that is not as apparent in other brood parasites. For example, an article in a North American journal solicited cowbird recipes (Schram, 1994)! In contrast, there is concern in Europe over the decline of the Common Cuckoo (Douglas et al., 2010). Birdwatchers and even some ornithologists in North America would likely rejoice if such declines occurred in the cowbird. This unfortunate attitude and scapegoating of the cowbird are detrimental to the conservation of endangered songbird species because it diverts attention from more important factors such as the anthropogenic habitat degradation underlying the declines of these species.
|
Ahmad M. 1990. Groundnut in the Dry Land Agriculture of Pothwar. Barani Agricultural Research and Development Project, PARC, Islamabad, Pakistan, p 43.
|
|
Ali S, Ripley SD. 1983. A Pictorial Guide to the Birds of the Indian Subcontinent. Oxford University Press, New York, p 35.
|
|
Ali S, Ripley SD. 1969. Handbook of the Birds of India and Paksitan. Vol. 3. Oxford University Press, Bombay, India.
|
|
Baker EC. 1921. Indian Game Birds, Pheasants, Bustard, Quail. Bombay Nat Hist Soc, London, UK.
|
|
Baker ECS. 1935. The Nidification of Birds of the Indian Empire. Taylor and Francis, London, p 4.
|
|
Beg AR, Baig MS, Ali Q, Khan CMA. 1985. Agro-ecological zonation of Potwar: a-wheat, b-maize. National Agricultural Research Centre, Islamabad, Pakistan Forest Institute, Peshawar and Soil Survey of Pakistan, Lahore.
|
|
Beg MA, Qureshi JI. 1972. Birds and their habitats in the cultivated areas of Lyallpur district and vicinity. Pakistan J Agric Sci, 9: 161–166.
|
|
Bump G, Bump JW. 1964. A study and review of the Black Francolin and the Grey Francolin. Special Scientific Report Wildlife (81): Bureau of Sport Fisheries and Wildlife, Washington D.C., USA 81: 53–57.
|
|
Burnham KP, Anderson DR, Laake JL. 1980. Estimation of density from line transects sampling of biological populations. Wildl Monogr, 72: 1–202.
|
|
Chaudhry AA, Bhatti MN. 1992. Biology of grey francolin (Francolinus pondicerianus) in the central Punjab plains. Abstract Proc. 12th Pakistan Congress of Zoology, Lahore, pp 161–162.
|
|
Clark A. 1901. Sport in the Low-country of Ceylon. Tisara Prakasakayo, Dehiwela.
|
|
del Hoyo J, Elliott A, Sargatal J (eds). 1994. Handbook of the Birds of the World. Volume 2. New World Vultures to Guineafowl. Lynx Edicions, Barcelona, Spain, pp 412–567.
|
|
Faruqui SA, Bump G, Nanda PC, Christensen GC. 1960. A study of the seasonal food of black francolin {Francolinus francolinus (Linnaeus)}, the gray francolin {Francolinus pondicerianus (Gmelin)} and the common sandgrouse {Pterocles exustus (Temminck)} in India and Pakistan. J Bombay Nat Hist Soc, 57: 354–361.
|
|
Gould J. 1966. Birds of Europe. Methuen, London, pp 49–50.
|
|
Islam K. 1999. Erckel's francolin (Francolinus erckelii), black francolin (Francolinus francolinus), and grey francolin (Francolinus pondicerianus). In: Poole A, Gill F (eds) The Birds of North America. The Birds of North America, Inc., Philadelphia, pp 394–396.
|
|
Javed S, Qureshi Q, Rahmani AR. 1999. Conservation status and distribution of swamp francolin in India. J Bombay Nat Hist Soc, 96: 16–23.
|
|
Johnsgard PA. 1973. Grouse and quails of North America. University of Nebraska, Lincoln, p 553.
|
|
Kaiser W. 1998. Autumn winter habitat used by radio-tagged grey partridge (perdix perdix) in north Bavaria. Game Wildlife Sci, 15: 471–479.
|
|
Kamal S. 2000. Population status, distribution pattern and habitat of grey partridge (Francolinus pondicerianus) in district Mardan. Dissertations, Pakistan Forest Institute, Peshwar, p 50.
|
|
Khan WA. 2010. Studies on the comparative ecology of the south Persian black francolin, Francolinus francolinus henrici, and northern grey partridge, Francolinus pondicerianus interpositus, in Lal Suhanra National Park, Bahawalpur, Punjab, Pakistan. Dissertations, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi, p 218.
|
|
Long JL. 1981. Introduced Birds of the World. David and Charles, London, p 89.
|
|
Mahmood K, Ahmad T, Khan A, Mahmood A, Mahmood W. 1997. Some notes on avifauna of Mangla reservoir, AJK, Pakistan. Pak J Ornith, 1: 1–2.
|
|
Mahmood S, Mahmood T, Rais M, Qureshi IZ, Nadeem MS. 2010. Comparative study on population and habitat of the grey and black francolins; Francolinus pondicerianus and Francolinus francolinus in Lehri Nature Park, Punjab, Pakistan. Podoces, 5(1): 42–53.
|
|
Mann MA, Chaudhry AA. 2000. Francolins in irrigated forest plantations and sub-mountainous tract of the Punjab, Pakistan. Pak Veterin J, 20(3): 118–122.
|
|
Mc Crow VP. 1982. Grey partridge habitat use and nesting biology in north central Iowa. Dissertations, Iowa State University, Ames, p 239.
|
|
Mian A, Ghani UI. 2007. Macrobiota variation under human protection in desert ecosystem of Cholistan, Pakistan. J Human Ecol, 21 (3): 163–172.
|
|
Mian A, Wajid MM. 1994. Food of grey francolin (Francolinus pondicerianus) in Layyah District (southern Punjab, Pakistan). Pakistan J Zool, 26: 185–187.
|
|
Mian A. 1995. Grey partridge demands intensive study. WPA News, 1: 2.
|
|
Nizami MI, Shafiq M, Rashid A, Aslam A. 2004. The soils and their agricultural development potential in Pothwar. WRRI and NARC, Islamabad, Pakistan, p 5–7.
|
|
Qureshi JI. 1972. Notes on the useful feeding activities of birds in field areas of Lyallpur. Pakistan J Agr Sci, 9 (3): 35–40.
|
|
Richard AF, Caroll JP, McGown JPK. 2002. Partridges, Quails, Francolins, Snowcocks, Guineafowl and Turkeys. Status survey and conservation. World Pheasant Associ. BirdLife International /SSC Partridges, Quails and Francolin specialist group. IUCN: The World Conservation Union. Gland, Switzerland, p 58.
|
|
Roberts TJ. 1991. The Birds of Pakistan, Non-passeriformes. Oxford University Press, New York, pp 232–233.
|
|
Shafiq MM. 2005. Wildlife Acts and Rules of Pakistan. unpublished report. Pakistan Forest Institute, Peshawar, p 297.
|
|
Sharma IK. 1983. The grey partridge (Francolinus pondicerianus) in the Rajasthan desert. Ann Arid Zone, 22: 117–120.
|
|
Ullah H. 1991. Studies on the biology, habitat, distribution pattern, and food of the grey partridge (Francolinus pondicerianus) in tehsil, Faisalabad. Dissertations, University of Agriculture, Faisalabad, p 182.
|
|
Waite HW. 1948. The birds of the Punjab salt range (Pakistan). J Bombay Nat Hist Soc, 48(1): 97–117.
|
|
Wijeyamohan S, Vandercone R, Santiapillai C. 2003. Observations on the grey francolin (Francolinus pondicerianus) in the vicinity of Giant's Tank, Sri Lanka. PQF News. 19: 11–14.
|
|
Yeatter RE. 1934. The Hungarian Partridge in Great Lakes Region. University of Michigan Press, Michigan, p 92.
|
Figures(3) / Tables(5)





 DownLoad:
DownLoad:


 Email Alerts
Email Alerts RSS Feeds
RSS Feeds