Ministry of Education, Key Laboratory for Biodiversity Sciences and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing 100875, China
Ultrasonic communication in vertebrates is attracting increasing research interest. To determine if ultrasonic vocalization is common in birds, we recorded their vocalizations with ultrasound detectors in the Dongzhai National Nature Reserve of Henan Province, China. We found varying degrees of high frequency components in the vocalizations of 14 species and in several of these species, the frequency of harmonics was up to the range of ultrasound. We suggest that more studies are required to determine whether the high frequency components in avian vocalizations have functions and what these functions are. In addition, the ability of birds to hear sounds in the high frequency range also requires re-examination.
Sound is widely used by animals for communication (Bradbury and Vehrencamp, 1998). Recently, a rapid expanding field in the study of animal communication is the use of ultrasonic (> 20 kHz) signals in terrestrial vertebrates (Arch and Narins, 2008). To this end, one of the more surprising groups shown to use ultrasound is amphibians (Narins and Feng, 2006), which, in the past, were thought to be unable to hear high frequency sounds (Pettigrew et al., 1981). However increasingly, evidence suggests that some frogs are capable of using ultrasonic vocalizations for communication (Feng et al., 2006; Arch et al., 2008; Arch et al., 2009).
However, the recent findings of ultrasonic communication in vertebrates (especially those in amphibians) suggest that communication approaches in the animal kingdom may be far more complex than previously thought. As birds are among those of which vocalizations are known to be most complicated (Dooling et al., 2002), it is therefore necessary and prudent to re-examine whether their vocalizations carry ultrasonic information. Normally, studies of avian vocalizations use standard recording equipment, with the upper limits of their frequency response seldom over 20 kHz. In our study, we used ultrasound detectors to conduct an investigation of the vocalizations of birds, aiming to detect whether ultrasonic vocalizations are common in birds.
Methods
All the avian sounds were recorded in the Dongzhai National Nature Reserve (31.95°N, 114.25°E; elevation 100–840 m) between January and March 2008, in June 2009 and December 2010. This reserve is located in the south of Henan Province of central China and within range of the Dabieshan Mountains. It is at the transitional region between the subtropical zone and the temperate zone and characterized by a rich avian diversity.
The sounds were recorded with either a Pettersson Bat Detector D980 (frequency range 10–200 kHz) or a D1000X (frequency range 5–235 kHz) (Pettersson Elektronik AB, Sweden). The detector D980 was connected to a laptop and sounds were recorded, using BatSound Pro software (Pettersson Elektronik AB, Sweden) and loaded onto the hard drive of the laptop. The D1000X is equipped with a built-in 16-bit recording system and stores sound as WAV files on a compact flash card and does therefore not require a laptop connection. We recorded bird vocalizations either in the field, or when the birds were temporarily caged. The target species was randomly chosen, depending on which species were found in the field or captured by mist nets. Distances to the birds varied from < 2 m for captive species to > 10 m for some species in the field. The sampling rates of the recordings were 100, 110, 192, 200, 300 or 384 kHz. The reason for the use of these many sampling frequencies was because we had no prior knowledge of the frequency range of the vocalization of a given species; thus they were usually selected in a tentative way. Spectrograms of the sounds were produced and checked for their frequency range with BatSound Pro. We selected spectrograms that were typical of the species to present in this paper.
Results
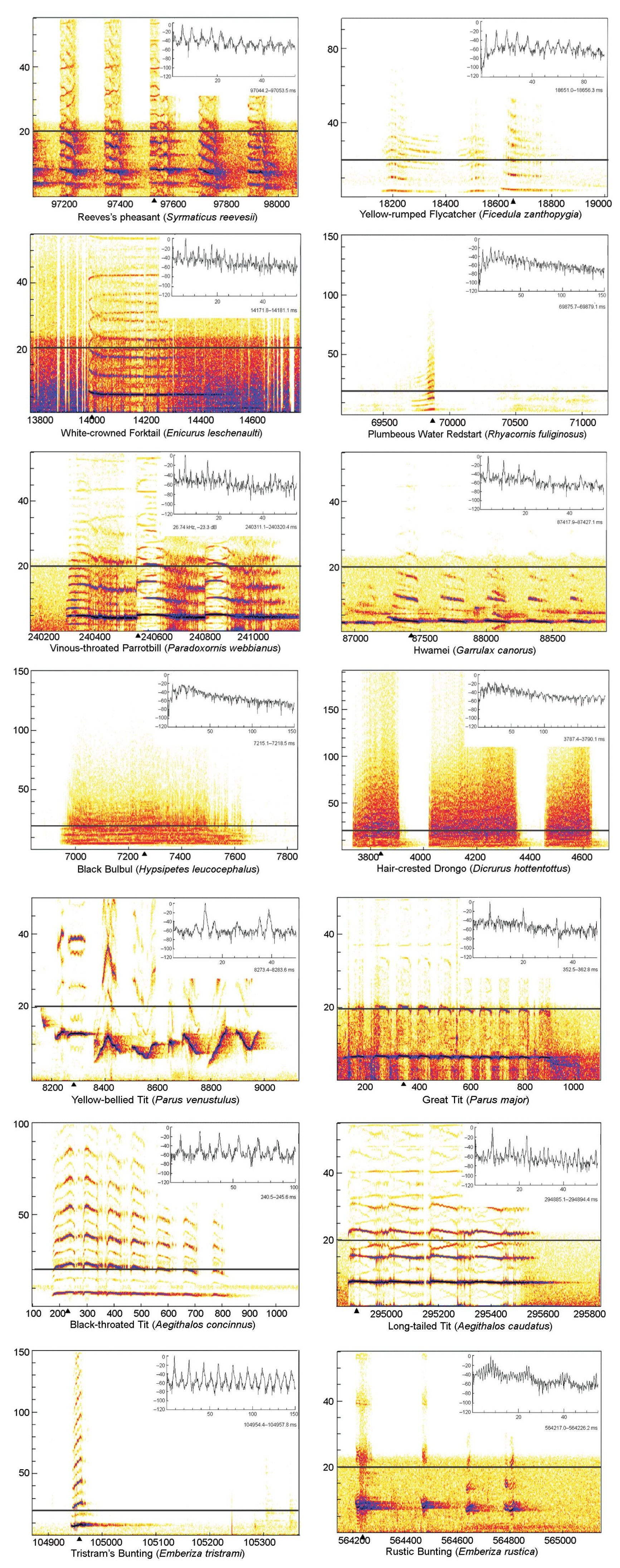
The vocalizations of 14 avian species were recorded (Table 1) and spectra of the selected vocaliztions are shown in Fig. 1.
Table
1.
Summary of the types of the avian vocalizations and recording sites
Figure
1.
Sound spectra and instantaneous amplitude spectra [insets, taken at indicated points (▲)] of the vocalizations of 14 avian species. The horizontal axis in the sound spectra represents the time (ms) and the vertical axis the frequency (kHz), while in the insets, the horizontal axis represents frequency (kHz) and the vertical axis amplitude (dB). The horizontal lines in the panels separate the ultrasonic and audible components of the vocalizations.
No bird was detected to produce pure ultrasonic calls. However, each bird vocaliztion contained more or less high frequency harmonics which approached or exceeded the audible limit of humans (20 kHz). The frequency of the harmonics of the Black-throated Tit and the Yellow-bellied Tit reached 40 kHz. It is noteworthy that despite a decreasing trend, the ultrasound components in some species, such as the White-crowned Forktail, Black-throated Tit, Long-tailed Tit and Yellow-bellied Tit, still carried significant energy, which was somewhat comparable to that of the fundamental components in their vocalizations (Fig. 1).
Discussion
Batsound detectors D980 and D1000X were able to record sounds with the highest frequency up to 200 kHz. During the study, we adopted sampling rates of at least 100 kHz, making the Nyquist frequencies at least 50 kHz, far exceeding the frequency range of human hearing. Therefore both the equipment and the sampling rates enabled us to detect ultrasonic components in avian vocalizations if there were any. In the vocalizations of 14 avian species, we demonstrated that they contained varying degrees of high frequency components and in several species, the frequency of the harmonics reached the ultrasonic range. As the energy of sounds decreased with an increase in the distance between recorder and producer, one limitation of the study became apparent in that the sounds of half of the species were recorded in the field, thus the highest frequency of some of the vocalizations shown in Fig. 1 might be lower than they actually were due to the effect of distance. However, this potential error could not conceal the fact of the existence of high frequency components in avian vocalizations, since the actual frequency might be even higher than those shown.
Because the harmonics in some species carried non-neglectable energy, the question arises as to why they produce those high frequency sounds if they have no function. Higher frequency vocalizations may be used adaptively to enhance the signal-to-noise ratio (Arch and Narins, 2008). It has been shown that in the Concave-eared Torrent Frog (Amolops tormotus), ultrasound communication is adopted to avoid masking by the wideband background noise of local fast-flowing streams where the species lives (Feng et al., 2006). In birds, there is also evidence that they may adjust the frequency of their vocalization in adaptation response to a noisy environment. Take the Great Tits for example, they have a higher minimum frequency of their songs at noisy locations, presumably to prevent their songs from being masked to some extent by the predominantly low-frequency ambient noise (Slabbekoorn and Peet, 2003). Therefore, it is reasonable to speculate that communication by high frequency vocalization might be favored by natural selection for species living in noisy environments, such as the White-crowned Forktail and Plumbeous Water Redstart, which live near streams.
However, the properties of high-frequency sounds such as limitation in long-distance signaling, directionality and susceptibility to scattering, may limit their use in communication (Smith, 1979; Arch and Narins, 2008). One may also argue that the high frequency harmonics are just byproducts of the sound production mechanism (Roverud, 1989), or they could just be providing a significant component to the tonal quality (Thorpe and Griffin, 1962). Despite these arguments, high frequency signals have their advantages under certain circumstances. They do not spread far and are therefore hard for predators and competitors to detect. Selective pressure may therefore favor their use in the presence of predators (Wilson and Hare, 2004; Wilson and Hare, 2006) and in communication for interactions between individuals at close quarters (Smith, 1979; Arch and Narins, 2008). Studies of Richardson's Ground Squirrels (Spermophilus richardsonii) have demonstrated the use of high frequency sound communication for predator avoidance. The squirrels can produce "whisper" calls containing pure ultrasonic frequencies of around 50 kHz, to warn nearby conspecifics of potential danger (Wilson and Hare, 2004; Wilson and Hare, 2006). In the present study, the call of a Great Tit was recorded when it alarmed its partner of our access. In the field, this call was also often heard when there were aerial predators, such as hawks, passing by the area. Since it contained significant energy both in fundamental and high frequency components (Fig. 1), this kind of alarm call may be more efficient to transmit information of the threat from signalers to their close conspecifics, before being heard by the predators.
One study has suggested birds are capable of discriminating harmonic complex (Dooling et al., 2002). The songs of the Yellow-bellied Tit inspired another idea about the function of these high frequency harmonics: could the high frequency harmonics signal individual quality? The spectrogram of the Yellow-bellied Tit in our study was recorded from a captive individual when it was singing. In the field, we noticed such singing often occurred when Yellow-bellied Tits were in flocks and displayed to each other. We therefore suggest these complex harmonic components in birds may carry information related to individual quality.
Currently, we cannot rule out any of these and/or other possibilities for the function of high frequency harmonics in avian vocalizations. It is clear that additional studies are needed to determine the functions of these high-frequency components. For example, playbacks of avian vocalizations, of which the lower frequency components have been filtered out, should be used to test whether species that are capable of producing high frequency vocalizations are able to hear them. If they have the ability to hear high frequency sounds, further experiments should then be designed to test possible adaptive hypotheses such as predator avoidance and choice of mates. Besides, although previous studies using electrophysiological approaches concluded that the sensitive frequency of birds was between 1–5 kHz (Dooling, 1978), other studies found that the upper limit of their hearing is usually at 20 kHz (reached after conditioning) and cochlear potentials have been detected at 25–30 kHz by using greater intensities of sound (Schwartzkopff, 1955). We therefore agree with Narins et al. (2004) that a systematic re-examination of avian hearing ability is required.
The 14 avian species involved in our study were from eight families of Passeriforms and one family of Galliformes, suggesting such high frequency harmonics exist at least in the vocalizations of some avian lineages such as Paridae, Aegithalidae and Emberizidae. Due to the small sample of the species, however, it is hard for us to conclude in which groups of avian species these high frequency components are more prevalent. As a result, we encourage more studies to be conducted in this area to explore the prevalence of high frequency vocalization in avian communication.
Acknowledgements
This work was supported by the National Basic Research Program of China (No. 2007CB41 1606). We thank Jinshuo ZHANG, Junxian SHEN, Canwei XIA, Yang QIU, Jiajun YING for their valuable help with the work, as well as Yang LIU and anonymous reviewers for their comments and suggestions to our manuscript.
Arch V, Grafe T, Gridi-Papp M, Narins P. 2009. Pure ultrasonic communication in an endemic Bornean frog. PLoS One, 4(4): 5413.
Arch V, Grafe T, Narins P. 2008. Ultrasonic signalling by a Bornean frog. Biol Lett, 4(1): 19–22.
Arch V, Narins P. 2008. "Silent" signals: Selective forces acting on ultrasonic communication systems in terrestrial vertebrates. Anim Behav, 76(4): 1423–1428.
Ballentine B, Hyman J, Nowicki S. 2004. Vocal performance influences female response to male bird song: an experimental test. Behav Ecol, 15(1): 163–168.
Beason R. 2004. What Can Birds Hear? USDA National Wildlife Research Center-Staff Publications, p 78.
Beer C. 1979. Vocal communication between laughing gull parents and chicks. Behaviour, 70(1): 118–146.
Bradbury J, Vehrencamp S. 1998. Principles of Animal Communication. Sinauer Associates, Sunderland, MA.
Dooling R, Leek M, Gleich O, Dent M. 2002. Auditory temporal resolution in birds: Discrimination of harmonic complexes. J Acoust Soc Am, 112: 748–759.
Dooling R. 1978. Behavior and psychophysics of hearing in birds. J Acoust Soc Am, 64: S4.
Feng A, Narins P, Xu C, Lin W, Yu Z, Qiu Q, Xu Z, Shen J. 2006. Ultrasonic communication in frogs. Nature, 440(7082): 333–336.
Forstmeier W, Balsby T. 2002. Why mated dusky warblers sing so much: territory guarding and male quality announcement. Behaviour, 139(1): 89–111.
Fullard JH, Barclay RMR, Thomas DW. 1993. Echolocation in free-flying Atiu swiftlets (Aerodramus sawtelli). Biotropica, 25(3): 334–339.
Gentner T, Hulse S. 2000. Female European starling preference and choice for variation in conspecific male song. Anim Behav, 59(2): 443–458.
Griffin D. 1953. Acoustic orientation in the oil bird, Steatornis. Proc Natl Acad Sci USA, 39(8): 884–893.
Konishi M, Knudsen EI. 1979. The oilbird: hearing and echolocation. Science, 204(4391): 425–427.
Konishi M. 1970. Comparative neurophysiological studies of hearing and vocalizations in songbirds. J Comp Physiol A, 66(3): 257–272.
Leonard M, Fernandez N, Brown G. 1997. Parental calls and nestling behavior in tree swallows. Auk, 114(4): 668–672.
Loffredo C, Borgia G. 1986. Male courtship vocalizations as cues for mate choice in the satin bowerbird (Ptilonorhynchus violaceus). Auk, 103(1): 189–195.
Narins P, Feng A, Lin W, Schnitzler H, Denzinger A, Suthers R, Xu C. 2004. Old World frog and bird vocalizations contain prominent ultrasonic harmonics. J Acoust Soc Am, 115: 910–913.
Narins PM, Feng AS. 2006. Hearing and sound communication in amphibians: prologue and prognostication. In: Narins PM, Feng AS, Fay RR, Popper AN (eds) Hearing and Sound Communication in Amphibians. Springer, New York, pp 1–11.
Pettigrew A, Anson M, Chung S. 1981. Hearing in the frog: a neurophysiological study of the auditory response in the midbrain. P Roy Soc Lond B Bio, 212(1189): 433–457.
Roverud R. 1989. Harmonic and frequency structure used for echolocation sound pattern recognition and distance information processing in the rufous horseshoe bat. J Comp Physiol A, 166(2): 251–255.
Schwartzkopff J. 1955. On the hearing of birds. Auk, 72(4): 340–347.
Slabbekoorn H, Peet M. 2003. Birds sing at a higher pitch in urban noise. Nature, 424(6946): 267–268.
Smith W. 1979. The study of ultrasonic communication. Am Zool, 19: 531–538.
Thorpe W, Griffin D. 1962. Ultrasonic Frequencies in Bird Song. Ibis, 104(2): 220–227.
Wilson D, Hare J. 2004. Animal communication: Ground squirrel uses ultrasonic alarms. Nature, 430(6999): 523.
Wilson DR, Hare JF. 2006. The adaptive utility of Richardson's ground squirrel (Spermophilus richardsonii) short-range ultrasonic alarm signals. Can J Zool, 84(9): 1322–1330.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds