Coastal Ecosystems Research Station of Yangtze River Estuary, Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Institute of Biodiversity Science, Fudan University, Shanghai 200433, China
2.
Chongming Dongtan National Nature Reserve, Chongming, Shanghai 202183, China
Variable and unpredictable food resources at stopover sites bring severe challenges to migrating shorebirds. Opportunistic foraging strategies, referring to shorebirds consuming prey in proportion to their availability, allow shorebirds to replenish fuel and nutrient reserves efficiently for continuing their migration. Chongming Dongtan, located in the Yangtze River estuary of eastern China, is the first major stopover site of shorebirds on the Chinese mainland during their northward migration. We investigated the diet of Great Knots (Calidris tenuirostris) at Chongming Dongtan during the spring stopovers of 2009 and 2010 through benthos sampling and dropping analysis. The benthos samples were categorized into gastropods, bivalves, polychaetes, crustaceans and insect larvae. Dropping analysis indicated that gastropods and bivalves constituted more than 70% of the diet of the Great Knot, with Assiminea violacea and Corbicula fluminea being the most frequently consumed. Chi-square tests indicated that for each prey category, there was no significant difference between the frequency of its occurrence in the benthos samples and dropping samples during the early stopover periods of 2009 and 2010 and during the late stopover periods of 2010. Although there was a statistically significant difference between the frequency of occurrence of prey in the total macrobenthos and in the droppings of the Great Knots during the late stopover period in 2009, the more abundant prey were more frequently consumed by the Great Knots. This suggests that Great Knots adopted an opportunistic foraging strategy during their stopover at Chongming Dongtan.
Most shorebirds need to replenish energy and nutrient reserves at one or more stopover sites during their long-distance migration. The quality of stopovers affects not only the success of migration but also the body condition when shorebirds arrive in the breeding grounds (Davis and Smith, 1998). Since the habitat quality at stopovers is highly dynamic, the availability of food resources for shorebirds is usually unpredictable. Thus, "opportunistic foraging", i.e. birds consuming prey in proportion to its availability, should be the optimal strategy in order to store fuel as soon as possible (Davis and Smith, 2001).
Millions of shorebirds stage at the coastal and estuarine wetlands in East Asia during their northward migration. The Great Knot (Calidris tenuirostris) is one of those long-distance migratory species along the East Asian-Australasian Flyway. In the breeding season, the adults and fledglings consume both berries and invertebrates, while the nestlings feed exclusively on insects and spiders (Higgins and Davies, 1996; Tomkovich, 1996). At stopover sites, bivalves constitute more than 60% (Wang and Qian, 1988) of the diet of Great Knots and 90% in their wintering grounds (Tulp and de Goeij, 1994). Red Knots (Calidris canutus), a species similar to the Great Knot, also prefer bivalves even though gastropods were available throughout the autumn stopover in the Wadden Sea (Dekinga and Piersma, 1993). These suggest that Great Knots might not be opportunistic foragers, but largely depend on the bivalves in non-breeding grounds and stopover sites. However, the temporary dynamics of their diet and the relationship between food use and food availability are still unknown.
In this paper, we demonstrate the diet composition and food resources of Great Knots in detail during their spring stopover at Chongming Dongtan, located in the Yangtze River estuary of eastern China. We also test whether Great Knots forage opportunistically or prefer bivalves during their stopover.
Study area
Chongming Island, with an area of 1200 km2, is the third largest island in China and the largest alluvial island in the world. Chongming Dongtan (31°29′N, 121°58′E) is located at the eastern end of the island and covers estuarine wetlands of 242 km2. Many birds depend on the wetlands during migration, wintering and breeding periods (Xu and Zhao, 2005). Chongming Dongtan is also the first major stopover site for shorebirds during their northward migration (Barter and Wang, 1990). Because of its importance for bird conservation, Chongming Dongtan has been designated as one of the wetlands of international importance (Ramsar sites) in 2002.
The estuarine wetlands at Chongming Dongtan are composed of bare mudflats in the low tidal zone and salt marshes in the middle and high tidal zones (Xu and Zhao, 2005). Most of the shorebirds, especially of the genus Calidris, inhabit and feed on plenty of macrobenthos in the intertidal zone during their stopover. The Great Knot is one of the most abundant Calidris species in the spring (Ma et al., 2004; Hui et al., 2009). Every year approximately 110000 Great Knots arrive at Chongming Dongtan in late March and leave before May (Barter, 2002).
Methods
Sampling
Macrobenthos and dropping samples were collected in 2009 and 2010 during the spring stopovers of Great Knots at Chongming Dongtan. Given the migratory phenology of the Great Knot, sampling was carried out twice each spring during the early (5 April in 2009 and 31 March in 2010) and late (18 April in 2009 and 19 April in 2010) stopover periods. All field work was carried out at low tide when the tidal flats were exposed and available for shorebirds to forage.
Binoculars (8 × 40) were used to search for the feeding area of Great Knots. After a flock of Great Knots left an area where they had foraged for at least half an hour, the time needed for food to pass through the digestive tract (Dekinga and Piersma, 1993), we collected droppings of Great Knots using medical spoons. Because Great Knots do not forage in fixed sites, our sampling also varied over the sampling periods. The samples were collected in sea bulrush marshes in both years, except for the late stopover period in 2009, when we took samples on the bare mudflats. Sometimes Great Knots foraged in mixed flocks with Dunlins (Calidris alpina), but the different size of droppings and footprints allow us to distinguish their droppings. About 30 droppings were sealed in a plastic bag as one sample.
Macrobenthos were sampled in the same feeding area where droppings were collected. To avoid the impact of bird foraging on benthos density, samples were taken on the mudflats where footprints and pecking marks of birds were absent. Ten benthos samples were randomly collected each time with metal corers (10 cm diameter) to a depth of 10 cm (Jing, 2005), with an interval of at least 10 m (González et al., 1996; D'Amico and Bala, 2004). The core samples were sieved on the spot through a 0.3 mm mesh and the macrobenthos were collected in plastic bottles. All samples were frozen at –20℃ until analyzed.
At the laboratory, the benthos samples were thawed in 75% ethanol. In each sample the prey items were identified to species level and counted under a binocular dissecting microscope (10 × 5) in a petri dish.
The composition of the diet of Great Knots was determined by faecal analysis. The dropping samples were dried at 60℃ and sieved through a 0.3 mm mesh to remove sediments (Dekinga and Piersma, 1993). We viewed dropping samples in a petri dish using the microscope (10 × 5) as mentioned earlier. Under the magnification used, one dropping sample could be scanned in about 30 scope fields. We quantified prey as the average per cent coverage of each field seen under the microscope, thus incorporating prey volume into this percentage measurement (Tsipoura and Burger, 1999). All representative hard prey remains were identified to the lowest possible taxon. Species identification was based on recognizable parts, such as hinges and shell fragments of bivalves, crab pincers and intact or broken shells of small gastropods.
Data analysis
Each prey category was summed up according to the sampling periods. The macrobenthos with a circumference of over 38 mm were excluded from analysis because food intake is limited by the gape width of the Great Knots (Tulp and de Goeij, 1994). By multiplying the sample size, the per cent coverage of each prey species in the droppings was converted into the frequency of its occurrence in the diet. Chi-square tests were used to determine whether the frequency of occurrence of prey in the droppings varied with the abundance of the same prey in the benthos samples, in 2009 and 2010. Crabs and polychaetes were excluded from the analysis because crabs moved rapidly and were difficult to detect while polychaetes could be completely digested by the Great Knots. Therefore, chi-square tests were based on the frequency of the occurrence of gastropods and bivalves, which occurred most frequently in the droppings of Great Knots (see Table 2). The level of statistical significance was set at 0.05.
Results
The potential food sources for Great Knots at Chongming Dongtan were composed of gastropods, bivalves, crustaceans, polychaetes and insect larvae (Table 1). The most abundant species were Assiminea violacea (Gastropoda), Corbicula fluminea (Bivalvia), Notomastus latericeus (Polychaeta), and Corophium volutator (Crustacea). Gastropods occurred at high density in both spring stopovers, while the density of the other three categories fluctuated. In spring 2009, bivalves increased sharply in the late stopover, while very few bivalves were collected in spring 2010. In contrast, the density of polychaetes changed in the two years and ranked second in 2010. There was a small amount of crustaceans in both years. Insect larvae occurred only accidentally.
Table
1.
Prey density (ind.·m–2) in the feeding areas of Great Knots at Chongming Dongtan in 2009 and 2010. All data are presented as average ± SD.
A total of 44 dropping samples (1100 droppings in total) were collected in two years. Dropping analysis indicated that the diet of Great Knots consisted of gastropods (Assiminea violacea), bivalves (Corbicula fluminea) and some small crabs (Ilyoplax deschampsi). The composition of the diet was different between the spring of 2009 and 2010 (Table 2). Gastropods dominated the diet almost during the entire stopover period, except for the late stopover in 2009, when bivalves became the main prey instead.
Table
2.
Cumulative percentage of prey items found in the droppings of Great Knots during spring stopovers at Chongming Dongtan
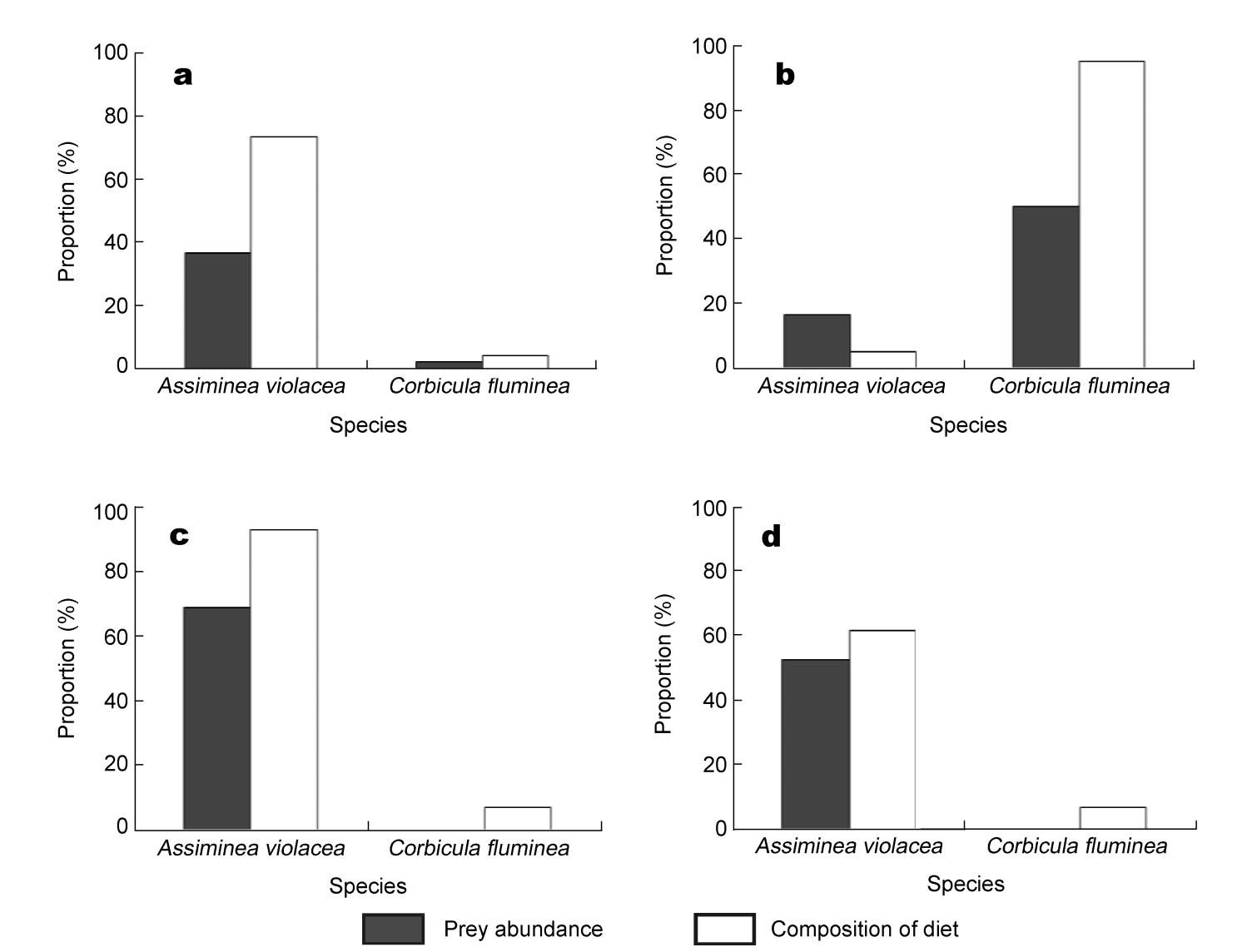
The data of Assiminea violacea and Corbicula fluminea was used in chi-square tests. There was no significant difference between the frequency of the occurrence of two prey in the diet of Great Knots and their abundance in benthos in both years except for the late stopover period of 2009 (Fig. 1; χearly, 20092 = 3.23, p > 0.05; χlate, 20092 = 13.19, p < 0.05; χearly, 20102 = 1.55, p > 0.05; χlate, 20102= 1.56, p > 0.05. df = 1 for all).
Figure
1.
Food availability and food use of Great Knots at Chongming Dongtan in four stopover periods. (a) early 2009; (b) late 2009; (c) early 2010; (d) late 2010.
Gastropods were the major benthos group at the feeding areas of Great Knots during the entire stopover period in 2010. However, in 2009, gastropods only dominated in the early stopover period and were later overtaken by bivalves. This might be caused by the change of feeding areas (from sea bulrush habitats to bare mudflats) in late 2009 when no foraging flocks were found in the former area. It also reflects a heterogeneous distribution of macrobenthos at Chongming Dongtan, with gastropods dominating the sea bulrush habitats while bivalves dominate the bare mudflats (Zhu et al., 2007).
Our results indicated that the frequency of occurrence of prey in the diet of Great Knots was not significantly different from their abundance, implying that the composition of the diet of Great Knots varied with the fluctuation of benthic fauna. This suggests that Great Knots adopt an opportunistic foraging strategy during their spring stopover at Chongming Dongtan.
Although dropping analysis indicated that the Ilyoplax deschampsi could be consumed by the Great Knots during spring stopovers, they were not detected in the macrobenthos samples. This species is likely to be underestimated as they can escape quickly during benthos sampling. In addition, polychaete worms could be fully digested by the Great Knots without leaving a trace in the droppings, which probably caused an undervaluation of the frequency of occurrence of such prey in the diet (Tulp and de Goeij, 1994; Barrett et al., 2007). Further studies, such as stable isotope analysis, might provide more information about the diet of Great Knots during stopovers.
The composition of the diet of Great Knots in this study is consistent with previous findings that bivalves, gastropods and crabs are prevalent invertebrates in stomach contents (Wang and Qian, 1988). However, we have established, for the first time, that gastropods are the most important prey for the Great Knots, especially during the early stopover period at Chongming Dongtan. In Roebuck Bay of Australia, the results of dropping analysis showed that 90% of the diet of Great Knots was made up of bivalves (Tulp and de Goeij, 1994). Such a contrast may arise due to the different macrobenthos composition between Chongming Dongtan and Roebuck Bay. In Roebuck Bay polychaetes are the most abundant with 70% of the benthic individuals, followed by bivalves (12.5%), while gastropods accounted for only 2.5% (Pepping et al., 1999). Because polychaetes can be digested without leaving a trace, it is not surprising that bivalves make up most of the diet in Roebuck Bay. Such variation in the composition of their diet also provides further evidence of the opportunistic foraging strategy of Great Knots.
Since shorebirds typically migrate across vast landscapes where wetlands are temporally and spatially dynamic (Skagen and Knopf, 1993; Farmer and Parent, 1997), it might be costly to discriminate between profitable and unprofitable prey (Davis and Smith, 2001). Consequently, the flexibility of an opportunistic foraging strategy enables shorebirds to replenish energy and nutrient reserves rapidly, which is important for them to complete their migratory flights and arrive on the breeding grounds in good condition as well (Davis and Smith, 1998).
Acknowledgements
This study was financially supported by the National Natural Science Foundation of China (Grant No. 30670269, 31071939). We thank the Chongming Dongtan National Nature Reserve for facilitating our fieldwork. We also thank Yong Wang, Jimmy Choi and two anonymous reviewers for their comments and suggestions on the original manuscript.
Barrett RT, Camphuysen CJ, Anker-Nilssen T, Chardine JW, Furness RW, Garthe S, Hüppop O, Leopold MF, Montevecchi WA, Veit RR. 2007. Diet studies of seabirds: a review and recommendations. ICES J Mar Sci, 64: 1675–1691.
Barter M, Wang TH. 1990. Can waders fly non-stop from Australia to China? Stilt, 17: 36–39.
Barter M. 2002. Shorebirds of the Yellow Sea: importance, threats and conservation status. Wetlands International Global Series 9. International Wader Studies 12. Canberra, Australia.
D'Amico VL, Bala LO. 2004. Diet of the Two-banded Plover in Caleta Valdés, Península Valdés, Argentina. Wader Study Group Bull, 103: 85–87.
Davis CA, Smith LM. 1998. Ecology and management of migrant shorebirds in the Playa Lakes Region of Texas. Wildl Monogr, 140: 1–45.
Davis CA, Smith LM. 2001. Foraging strategies and niche dynamics of coexisting shorebirds at stopover sites in the Southern Great Plains. Auk, 118: 484–495.
Dekinga A, Piersma T. 1993. Reconstructing diet composition on the basis of faeces in a mollusc-eating wader, the knot Calidris canutus. Bird Study, 40: 144–156.
Farmer AH, Parent AH. 1997. Effects of the landscape on shorebird movements at spring migration stopovers. Condor, 99: 698–707.
González PM, Piersma T, Verkuil Y. 1996. Food, feeding, and refuelling of Red Knots during northward migration at San Antonio Oeste, Rio Negro, Argentina. J Field Ornithol, 67: 575–591.
Higgins PJ, Davies JN. 1996. Handbook of Australian, New Zealand and Antarctic Birds. Volume 3. Royal Australasian Ornithologists Union, Oxford University Press, Melbourne, pp 214–224.
Hui X, Ma Q, Xiangyu JG, Cai ZY, Song GX, Yuan X, Ma ZJ. 2009. Migration routes of shorebirds across Chongming Dongtan based on recovery of bird banding. Chinese J Zool, 44(3): 23–29. (in Chinese with English abstract)
Jing K. 2005. Stopover Ecology of Shorebirds in Chongming Dongtan. Dissertation. Fudan University, Shanghai.
Ma ZJ, Li B, Zhao B, Jing K, Tang SM, Chen JK. 2004. Are artificial wetlands good alternatives to natural wetlands for waterbirds? A case study on Chongming Island, China. Biodivers Conserv, 13: 333–350.
Pepping M, Piersma T, Pearson G, Lavaleye M. 1999. Intertidal sediments and benthic animals of Roebuck Bay Western Australia. NIOZ-report, 1999-3, pp 195–196.
Tomkovitch PS. 1996. A third report on the biology of the Great Knot Calidris tenuirostris, on the breeding grounds. Stilt, 28: 43–45.
Tsipoura N, Burger J. 1999. Shorebird diet during spring migration stopover on Delaware Bay. Condor, 101: 635–644.
Tulp I, de Goeij P. 1994. Evaluating wader habitats in Roebuck Bay (north-western Australia) as a springboard for northbound migration in waders, with a focus on Great Knots. Emu, 94: 78–95.
Wang TH, Qian GZ. 1988. Shorebirds in the Yangtze River Estuary and Hangzhou Bay. East China Normal University Press, Shanghai. (in Chinese)
Xu HF, Zhao YL. 2005. Comprehensive Surveys in Chongming Dongtan Nature Reserve for Migratory Birds, Shanghai. China Forestry Publishing House, Beijing. (in Chinese)
Zhu J, Jing K, Gan XJ, Ma ZJ. 2007. Food supply in intertidal area for shorebirds during stopover at Chongming Dongtan, China. Acta Ecol Sinica, 27: 2149–2159.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds