Figure
1.
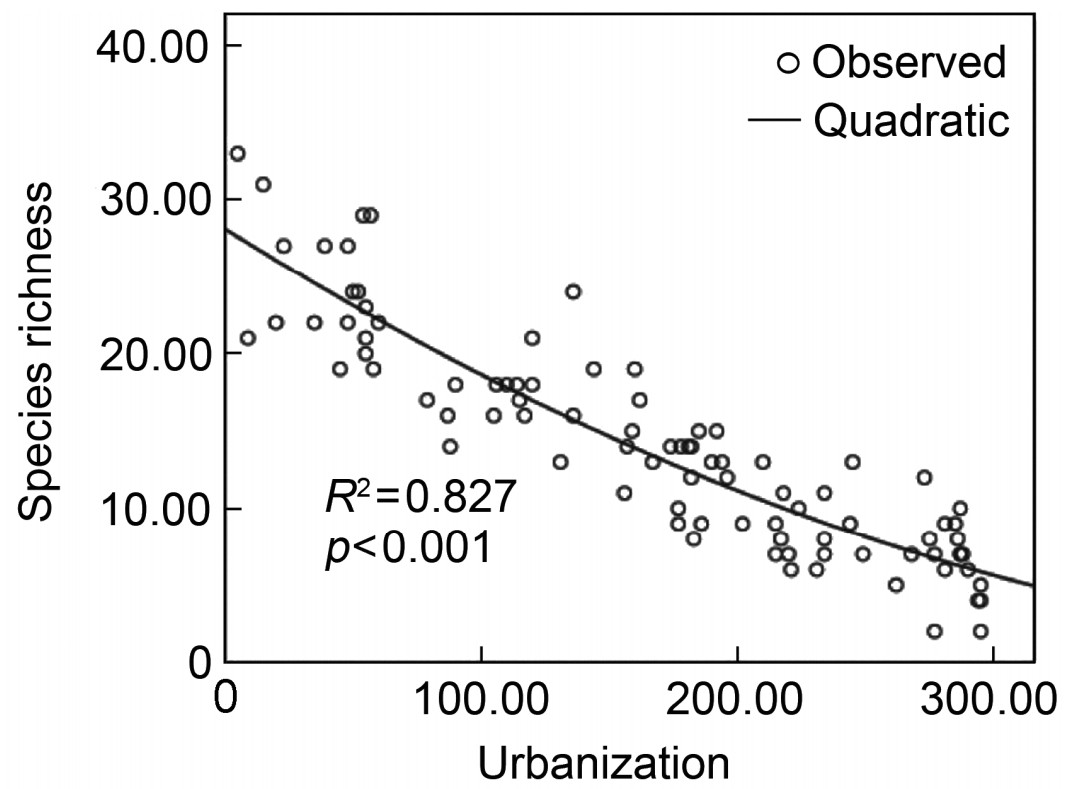
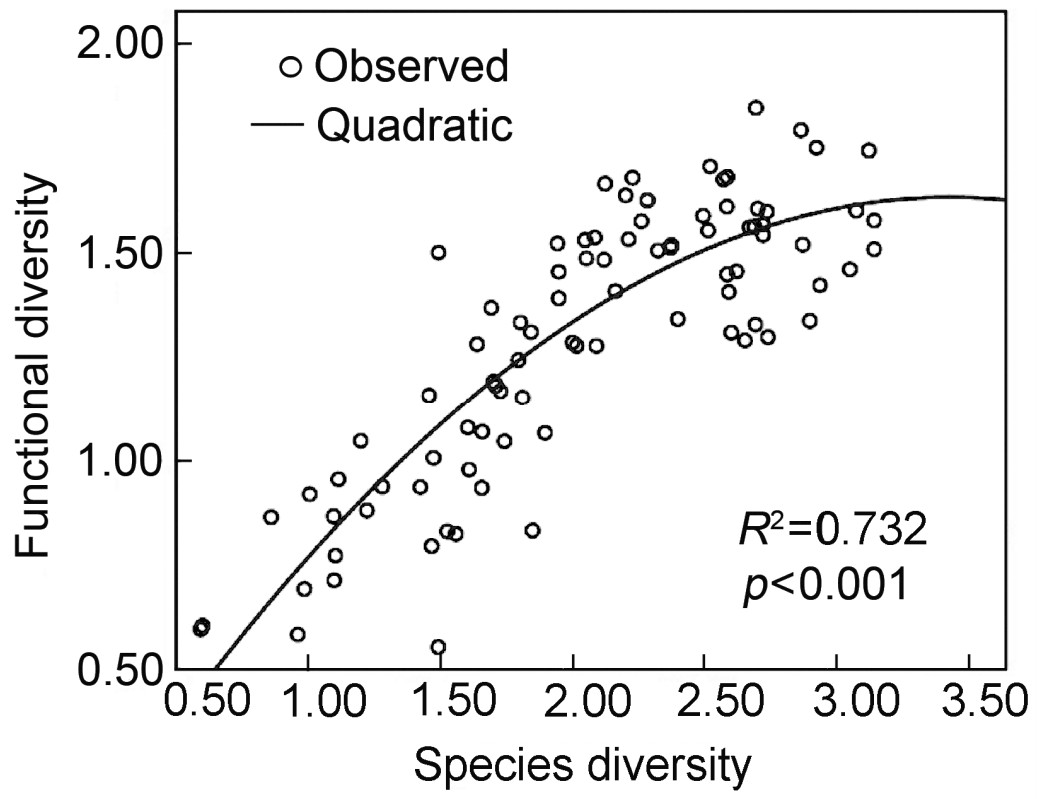
Quadratic curve fitted of species richness versus synthetic urbanization index
| Citation: |
Peng LI, Ping DING, Kenneth J. FEELEY, Jingcheng ZHANG, Pingping JIANG. 2010: Patterns of species diversity and functional diversity of breeding birds in Hangzhou across an urbanization gradient. Avian Research, 1(1): 1-8. DOI: 10.5122/cbirds.2009.0020

|
Given the rapid rise in human population and increasing urbanization,it is important to understand their potential impacts on biodiversity. From March 2007 to August 2007, we conducted bird surveys in 90 strip transects,each 3 km long and 100 m wide,along a gradient of urbanization in Hangzhou,China. This gradient spanned a range of urbanization levels including urban areas,rural-urban continuum areas,farming areas,mixed forest/farming areas and forested areas. We recorded 96 breeding bird species and classified them into nine functional groups based on nesting requirements. The nine functional groups consisted of canopy nesters,shrub nesters,canopy/shrub nesters,natural cavity nesters,building nesters,natural cavity/building nesters,ground nesters,water surface nesters and parasitic nesters. Species and functional diversities were estimated based on the Shannon-Wiener index. Environmental data of each transect as human disturbance,vegetation cover and building index were also measured,and a synthetic urbanization index of each transect was introduced based on these data. We used regression analyses to model the relationship of species abundance,species diversity,functional abundance and functional diversity with this synthetic index. The results show that urbanization significantly reduces species richness,species diversity,functional richness and functional diversity,but the specific patterns differed. The relationship between species abundance/species diversity and urbanization is linear. In contrast,the relationship between functional diversity and urbanization was quadratic. In other words,with increased urbanization,functional diversity declined only slightly at first but then dropped at an accelerating rate. This implies that,although moderate urbanization reduces species diversity of breeding birds,it affects functional diversity of breeding birds only slightly in Hangzhou. The regression analysis of species diversity and functional diversity suggests a quadratic relationship between species diversity and functional diversity,i.e.,a linear relationship between species diversity and functional diversity can only exist at low diversity levels across urbanization gradients and increasing species abundance does not lead to an increase in functional diversity at the highest diversity levels.
Global biodiversity is being rapidly lost (Díaz and Cabido, 2001) and urbanization is regarded as one of the primary factors driving the decline (Garden et al., 2006). Many researchers have investigated the potential effect of increasing urbanization on species diversity and especially the diversity of avian communities.
Most researchers believe that urbanization generally decreases species richness and species diversity (Emlen, 1974; Beissinger and Osborne, 1982; DeGraaf and Wentworth, 1986; Rosenberg et al., 1987; Rottenborn, 1999; Melles et al., 2003; Lee et al., 2004). However, other researchers have argued that, because of new food and habitats introduced by urbanization (Blair, 1996), bird communities may actually reach their highest richness and diversity in moderately urbanized areas (Blair, 1996, 2004; Crooks et al., 2004).
However, these previous studies have focused primarily on species richness or species diversity which is only one component of biodiversity (Díaz and Cabido, 2001; Moretti et al., 2009). Biodiversity also includes genetic, phylogenetic and functional diversity (Tilman, 2001). Moreover, increasing numbers of researchers have agreed that it is functional trait, rather than taxonomic identity, that has the greatest effect on ecosystem processes (Díaz and Cabido, 2001; Hooper et al., 2002; McGill et al., 2006; Pavao-Zuckerman and Coleman, 2007; Knapp et al., 2008; Moretti et al., 2009). McGill et al. (2006) go as far as to suggest that functional traits are the key to rebuilding community ecology.
Therefore, in this study, we investigated breeding bird communities along an urbanization gradient and looked at the impact of increasing urbanization on one key functional trait — nesting requirement. We explored how urbanization affects species diversity and functional diversity respectively and whether the patterns differ or not. Specifically we tested the hypotheses that: 1) there are negative correlations between species richness, diversity and urbanization; 2) functional species and diversity are negatively correlated with urbanization; and 3) the form of the relationship between species diversity and functional diversity is asymptotic. The findings of this study will help guide the conservation of bird communities in an increasingly urban landscape.
The study was conducted in the Hangzhou region (29°12′–30°35′N, 118°12′–120°43′E), situated in southeastern China with an area of 16596 km2. The western part of the region is hilly while the eastern portion is relatively flat. The hilly area accounts for about 66% of the total region, the flatter plain 26% and water 8% (Hangzhou Bureau of Statistics, http://www.hzstats.gov.cn). The highest point in the region is Qingliang Mountain, which reaches 1787 m a.s.l. and is located along the western border. Hangzhou has a subtropical monsoon climate with four distinct seasons. Its forest cover is approximately 11000 km2 (Hangzhou Bureau of Statistics, http://www.hzstats.gov.cn).
Birds were surveyed in 90 strip transects (3000 m × 100 m), placed along a gradient of urbanization including habitats suffering from different degrees of urbanization, including rural-urban continuum areas, farming areas, mixed forest/farming areas and forested areas. Surveys were conducted from March 2007 to August 2007 by two or three observers between 6:00–10:00 hours and 16:00–18:00 hours on clear days with little or no wind. Because we chose nesting requirement as the functional trait, only breeding birds seen or heard in transects were recorded and birds of passage were ignored.
We categorized all of the recorded bird species into nine discrete mutually-exclusive functional groups according to nesting requirements. The groups were: canopy nesters, shrub nesters, canopy/shrub nesters, natural cavity nesters, building nesters, natural cavity/building nesters, ground nesters, water surface nesters and parasitic nesters (Canterbury et al., 2000; Fernández-Juricic, 2000; Park and Lee, 2000; Lindsay et al., 2002; Lim and Sodhi, 2004; Palomino and Carrascal, 2006; Fontaine et al., 2007). Information about nesting requirements was obtained from observations when conducting the surveys and from published literatures (Cheng et al., 1978, 1979, 1997; Li et al., 1982; Zheng, 1985; Cheng et al., 1987, 1991, 1995; Zheng, 1995; Chen and Luo, 1998; Fu et al., 1998; Sun et al., 2002; Zheng and Zhang, 2002; Li et al., 2004).
Species diversity was calculated based on the Shannon-Wiener index:
|
|
(1) |
where H′ is the value of species diversity, S the total number of species in the community and Pi the proportion of the ith species in a sample.
Given different research objects and aims, there are various ways to measure functional diversity (Tilman, 2001). Generally, all these measures can be classified into two categories: discontinuous or continuous measures, based on the traits being discontinuous or continuous (Petchey and Gaston, 2006).
Because of the discontinuity trait used here (nesting requirement), we adopted a discontinuous measure which defines functional richness simply as the number of functional groups (Díaz and Cabido, 2001; Tilman, 2001; Naeem and Wright, 2003) and functional diversity as the Shannon-Wiener diversity of functional groups (Stevens et al., 2003; Petchey and Gaston, 2006), but where H′ is the value of functional diversity, S the total number of functional groups in the community and Pi the proportion of the ith functional groups in a sample.
Many variables have been used to characterize the degree of urbanization in previous studies, such as per cent vegetation cover (Rottenborn, 1999), distance to city center (Bolger et al, 1997; Wang et al, 2008), percentage of areas covered by building (Mills et al., 1989; Blair, 1996; Bolger et al., 1997; Lim and Sodhi, 2004; Palomino and Carrascal, 2006), percentage of paved areas (Palomino and Carrascal, 2006), the amount of pavement (Mills et al., 1989; Blair, 1996), environmental noise (Wang et al., 2008) and human population density (Lim and Sodhi, 2004). However, city landscapes are so complex that any single urbanization characteristic variable can only poorly reflect the true degree of urbanization. Therefore, in this study we used three different characteristic variables, i.e., human disturbance, vegetation cover and buildings. Details of these variables appear in Li et al. (2009). Furthermore, we combined these three variables using a synthetic urbanization index (SUI) (Chen et al., 2000), where SUI = (20 × human disturbance) + [(100 – (100 × vegetation cover)] + (100 × building index).
We used linear, quadratic, and cubic curve regression analyses to model the relationship of species richness, species diversity, functional richness and functional diversity with the synthetic urbanization index. In addition, we also used linear, quadratic, and cubic curve regression analyses to model the relationship of species diversity with functional diversity. All analyses were performed with SPSS (SPSS Inc., 2007).
Given our analyses, the best model of the relationship between species richness and the urbanization synthetic index turned out to be a quadratic curve (Fig. 1) as described by the equation: y = −0.105x + 9.887×10−5x2 + 28.122, R2 = 0.827 (F = 207.450, p < 0.001).
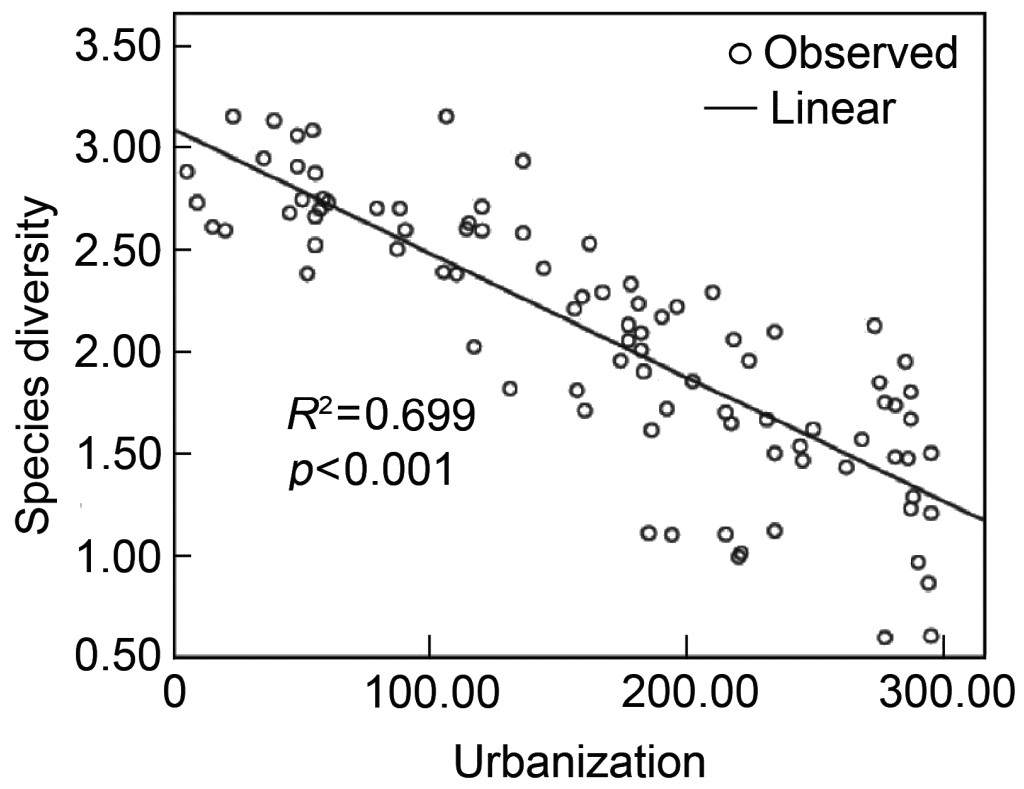
The best model of relationship between species diversity and the synthetic urbanization index is a straight line (Fig. 2) as described by the equation: y = −0.006x + 3.080, R2 = 0.699 (F = 204.733, p < 0.001).
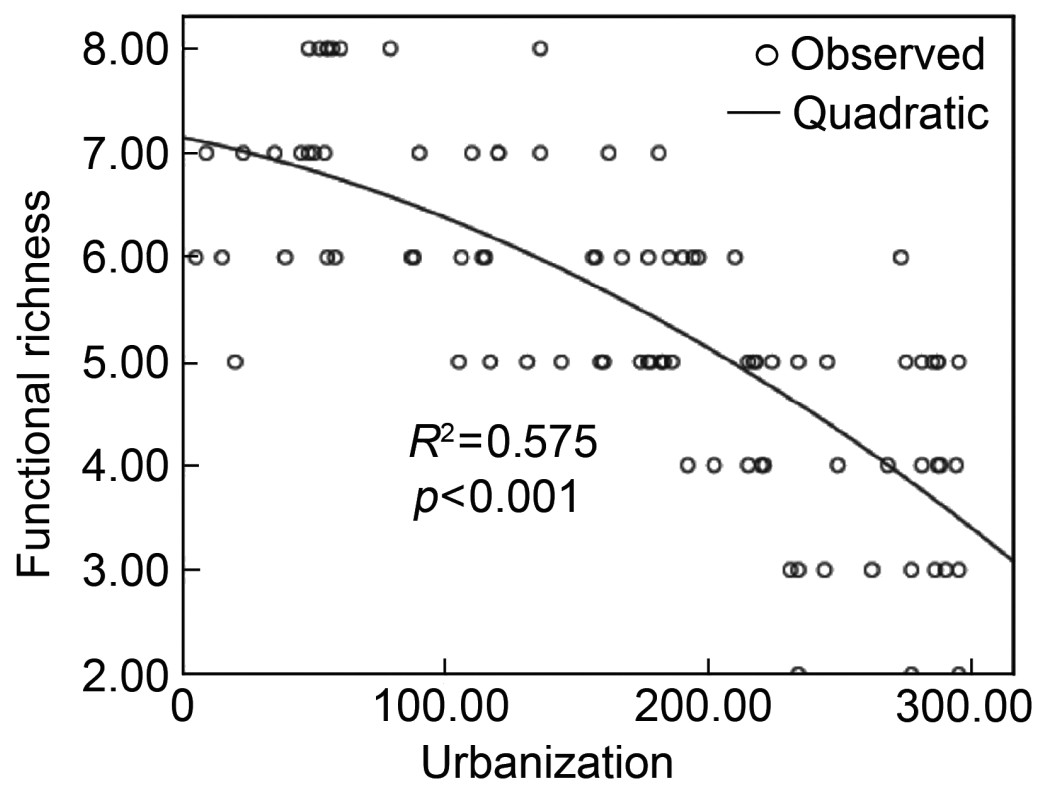
From our analyses, we conclude that the best model describing the relationship between functional richness and the synthetic urbanization index was also quadratic (Fig. 3) as described by the equation: y = −0.005x − 2.400×10−5x2 + 7.150, R2 = 0.575 (F = 58.862, p < 0.001).
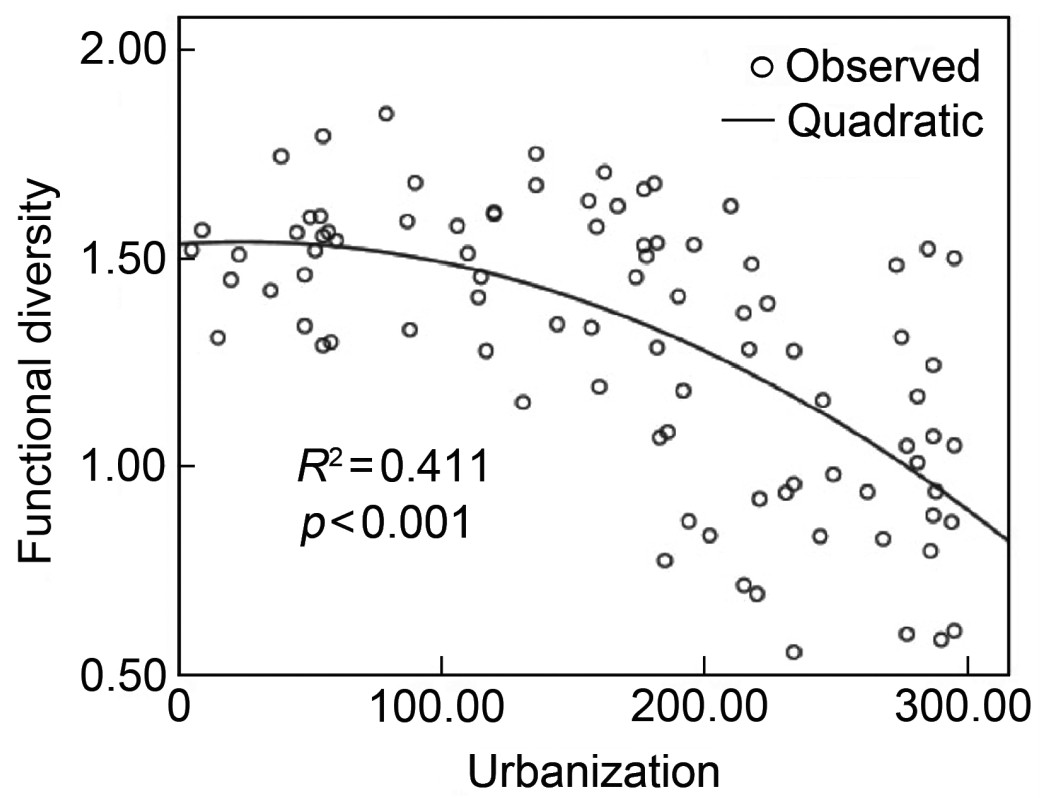
The best model of the relationship between functional diversity and the synthetic urbanization index was a quadratic curve (Fig. 4) as described by the equation: y = −8.498×10−6x2 + 1.534, R2 = 0.411 (F = 30.065, p < 0.001).
According to our analysis, the best model of the relationship between species diversity and functional diversity was also a quadratic curve (Fig. 5) as described by the equation: y = 1.003x − 0.146x2 − 0.092, R2 = 0.732 (F = 117.620, p < 0.001).
The effect of urbanization on bird species diversity has received a great deal of attention with most studies reporting, that because of changes in habitats, fragmentation and disturbance, urbanization reduces species richness and diversity (Emlen, 1974; Beissinger and Osborne, 1982; DeGraaf and Wentworth, 1986; Rosenberg et al., 1987; Rottenborn, 1999; Melles et al., 2003; Crooks et al., 2004; Lee et al., 2004). Lee et al. (2004) verified that urbanization can lead to a low level of primary ecosystem productivity, which in turn may cause a decline in diversity. Moreover, the research by Crooks et al. (2004) indicated that there are strong correlations between the size of fragmented areas introduced by urbanization, bird species abundance and diversity.
On the other hand, some researchers have contended that bird species richness and diversity should increase with urbanization because of the extra habitats and food sources made available (Mills et al., 1989; Blair, 1996, 1999, 2004; Crooks et al., 2004).
Our results indicate that in Hangzhou, both the breeding bird species richness and species diversity decreased with increasing urbanization and we found no evidence that bird species richness and diversity peaked in intermediate urbanized areas. The fact that diversity did not increase at an intermediate level of urbanization may reflect relative intense urbanization and human disturbance in this region.
Ricklefs (2001) suggested that biodiversity could be understood as a niche relationship, i.e., biodiversity is related to the abundance of niches, niche breadth and degree of overlap of niches in an area. For example, in Ontario, southern Canada, bat species have similar phenotypes because of their simpler food resources. They are all miniature insectivorous with similar roles in ecosystem functioning. Meanwhile, Cameroon, located in tropical western Africa, offers a greater diversity of food resources and the different bat species are functioning in various ecosystem roles such as frugivores, nectavores, piscivores and predators as well as insectivores (Ricklefs, 2001).
Stevens et al. (2003) also discovered that richness, diversity and dominance of functional groups all decreased along latitudinal gradients because food resources are reduced at higher latitudes.
Furthermore, research of fish communities in the Rio Tamesi, a river in eastern Mexico, has indicated that the number of fish species and functional groups increases downstream in the river. This is believed to be due to the fact that food resources increase and diversify with the widening of the river (Ricklefs, 2001).
Our results showed that bird functional richness and diversity declined with urbanization. Based on our previous studies, we hypothesize that this is due to a decrease in nesting sites with greater urbanization and human disturbance.
We found that functional diversity did not drop markedly at the lower level of urbanization. This may indicate that at the earliest stages of urbanization there is a low level of natural habitat destruction/disturbance and any disturbance may be offset by new nesting sites offered by buildings or man-made structures. However, with increasing urbanization, more and more natural habitats are transformed into artificial sites and although the absolute number of nesting sites may increase or remain stable, the variety of nesting sites declines as habitats become homogenized. At the highest levels of urbanization, even more natural habitats are lost and the degree of anthropogenic effects is enhanced, resulting in a dramatic decline in the functional diversity of breeding birds.
The form of the relationship between species diversity and functional diversity is often modeled as asymptotic (Naeem, 2002). All the same, patterns vary with different number of traits or functional groups (Petchey and Gaston, 2002). Petchey and Gaston (2002) thought that with an increasing number of traits, the relationship between species diversity and functional diversity would tend to be linear; while with an increase in the number of functional groups, a discrete relationship would transform into a continuous one. Of course, different measures of functional diversity would affect the specific shape of the relationship (Petchey and Gaston, 2006).
Our results illustrate that, although both species diversity and functional diversity declined with urbanization, the patterns were different (Figs. 2 and 4). Species diversity declined linearly with urbanization, while functional diversity declined slowly at early stages of urbanization and decreased rapidly during later stages.
According to the regression model of the relationship between species and functional diversity (Fig. 5), we found that the relationship was quadratic. It is similar to the results obtained by Stevens et al. (2003) in their study of functional diversity of bats in America across latitudinal gradients. Hooper et al. (2002) also indicated that species diversity was highly correlated with functional diversity at the lower levels of biodiversity, yet functional diversity could not catch up with species diversity with rising biodiversity.
We found that breeding bird functional diversity declined slightly at the beginning of urbanization. That might be because there were sufficient functional groups in the community and sufficient species in each group on this level, so that a slight loss of individuals or species would not reduce the number of functional groups or functional diversity. This was found to be the case in a study by Petchey and Gaston (2006). They found that the greater the number of functional differences in a community, the greater its functional redundancy. All the same, with urbanization, more and more individuals or species are lost to the point that functional richness declines. In heavily urbanized areas, there were few species in each group and the loss of each species would likely cause a corresponding loss of a functional group. Therefore, the relationship of species diversity and functional diversity tended to be linear at high levels of urbanization.
We come to the conclusion that moderate urbanization decreases species diversity of breeding birds but, so far, has not markedly affected functional diversity of breeding birds in Hangzhou, while heavy urbanization caused rapid declines in both species and functional diversity. The loss of functional diversity has especially important implications since it may potentially lead to the loss of ecosystem functions or trophic effects on other animal groups. Therefore, we suggest that attempts should be made to maintain future urbanization at a moderate level. We also suggest that more attention be paid to functional traits, rather than taxonomic identities, in community ecology and conservation studies (McGill et al., 2006).
We thank Bicheng Li, Guirong Xia, Chris Wood and Meng Zhang for their help in field surveys. This study was supported by the National Natural Science Foundation of China (Grant No. 30670344) and the Hangzhou Municipal Bureau of Forestry and Water Resources.
|
Beissinger SR, Osborne DR. 1982. Effects of urbanization on avian community organization. Condor, 8:75–83
|
|
Chen SH, Ding P, Zheng GM, Zhuge Y. 2000. Impacts of urbanization on the wetland water-bird communities in Hangzhou. Zool Res, 21(4):279–285 (in Chinese)
|
|
Chen FG, Luo SY. 1998. Fauna Sinica Aves Vol.9. Science Press, Beijing (in Chinese)
|
|
Cheng TH. 1978. Fauna Sinica Aves Vol.4. Science Press, Beijing (in Chinese)
|
|
Cheng TH. 1979. Fauna Sinica Aves Vol.2. Science Press, Beijing (in Chinese)
|
|
Cheng TH, Long ZY, Zheng BF. 1987. Fauna Sinica Aves Vol.11. Science Press, Beijing (in Chinese)
|
|
Cheng TH, Xian YH, Guan GX. 1991. Fauna Sinica Aves Vol.6. Science Press, Beijing (in Chinese)
|
|
Cheng TH, Long ZY, Lu TC. 1995. Fauna Sinica Aves Vol.10. Science Press, Beijing (in Chinese)
|
|
Cheng TH. 1997. Fauna Sinica Aves Vol.1. Science Press, Beijing (in Chinese)
|
|
Fu TS, Song YJ, Gao W. 1998. Fauna Sinica Aves Vol.14. Science Press, Beijing (in Chinese)
|
|
Hooper DU, Solan M, Symstad A, Díaz S, Gessner MO, Buchmann N, Degrange V, Grime P, Hulot F, Mermillod-blondin F, Roy J, Spehn E, van Peer L. 2002. Species diversity, functional diversity and ecosystem functioning. In: Loreau M, Naeem S, Inchausti P (eds) Biodiversity and Ecosystem Functioning: Syntheses and Perspectives. Oxford University Press, Oxford, pp 195–208
|
|
Li GY, Zheng BL, Liu GZ. 1982. Fauna Sinica Aves Vol.13. Science Press, Beijing (in Chinese)
|
|
Li PF, Li W, Zhou W, Wang CF, Cai HY. 2004. Ecological habits of bird of hawk family in Baishilazi region. J Liaoning Forest Sci Technol, 3:5–8 (in Chinese)
|
|
Melles S, Glenn S, Marti K. 2003. Urban bird diversity and landscape complexity: Species-environment associations along a multiscale habitat gradient. Conserv Ecol, 7(1):5
|
|
Moretti M, Bello F, Roberts SPM, Potts SG. 2009. Taxonomical vs. functional responses of bee communities to fire in two contrasting climatic regions. J Anim Ecol, 78:98–108
|
|
Naeem S. 2002. Autotrophic–heterotrophic interactions and their impacts on biodiversity and ecosystem functioning. In: Kinzig AP, Pacala SW, Tilman D (eds) The Functional Consequences of Biodiversity. Princeton University Press, Princeton, NJ, pp 120–150
|
|
Ricklefs RE. 2001. The Economy of Nature, 5th edn. W.H. Freeman and Company, New York
|
|
Rosenberg KV, Terrill SB, Rosenberg GH, 1987. Value of suburban habitats to desert riparian birds. Wilson Bull, 99: 643–654.
|
|
Sun MY, Li KQ, Zhu JJ, Kao CC, Sun XJ. Zhou ZS. 2002. Reproduction habits of three species of woodpeckers and their prey on insects. Forest Pest Dis, 21(2):12–14 (in Chinese)
|
|
Tilman D. 2001. Functional diversity. In: Levin SA (ed) Encyclopedia of Biodiversity, Vol 3. Academic Press, San Diego, CA, pp 109–120
|
|
Wang YP, Chen SH, Jing PP, Ding P. 2008. Black-billed Magpies (Pica pica) adjust nest characteristics to adapt to urbanization in Hangzhou, China. Can J Zool, 86:868–874
|
|
Zheng BL. 1985. Fauna Sinica Aves Vol.8. Science Press, Beijing (in Chinese)
|
|
Zheng GM. 1995. Ornithology. Beijing Normal University Press, Beijing (in Chinese)
|
|
Zheng GM, Zhang CZ. 2002. Birds in China. China Forestry Publishing House, Beijing (in Chinese)
|
Figures(5)





 DownLoad:
DownLoad:
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds