Adam, I., Maxwell, A., Rößler, H., Hansen, E.B., Vellema, M., Brewer, J., et al., 2021. One-to-one innervation of vocal muscles allows precise control of birdsong. Curr. Biol. 31, 3115–3124.
|
Akutagawa, E., Konishi, M., 2010. New brain pathways found in the vocal control system of a songbird. J. Comp. Neurol. 518, 3086–3100.
|
Al-Yaari, M., Yamada, R., Kuba, H., 2020. Excitatory–inhibitory synaptic coupling in avian nucleus magnocellularis. J. Neurosci. 40, 619–631.
|
Alger, S.J., Larget, B.R., Riters, L.V., 2016. A novel statistical method for behaviour sequence analysis and its application to birdsong. Anim. Behav. 116, 181–193.
|
Alonso, R., Goller, F., Mindlin, G.B., 2014. Motor control of sound frequency in birdsong involves the interaction between air sac pressure and labial tension. Phys. Rev. E. 89, 032706.
|
Alvarez-Borda, B., Nottebohm, F., 2002. Gonads and singing play separate, additive roles in new neuron recruitment in adult canary brain. J. Neurosci. 22, 8684–8690.
|
Aronowitz, J.V., Perez, A., O'Brien, C., Aziz, S., Rodriguez, E., Wasner, K., et al., 2021. Unilateral vocal nerve resection alters neurogenesis in the avian song system in a region-specific manner. PLoS One 16, e0256709.
|
Bailey, D.J., Rosebush, J.C., Wade, J., 2002. The hippocampus and caudomedial neostriatum show selective responsiveness to conspecific song in the female zebra finch. J. Neurobiol. 52, 43–51.
|
Ball, G.F., 1994. Neurochemical specializations associated with vocal learning and production in songbirds and budgerigars. Brain Behav. Evol. 44, 234–246.
|
Ballentine, B., Hyman, J., Nowicki, S., 2004. Vocal performance influences female response to male bird song: an experimental test. Behav. Ecol. 15, 163–168.
|
Barnea, A., Mishal, A., Nottebohm, F., 2006. Social and spatial changes induce multiple survival regimes for new neurons in two regions of the adult brain: an anatomical representation of time? Behav. Brain Res. 167, 63–74.
|
Barnea, A., Pravosudov, V., 2011. Birds as a model to study adult neurogenesis: bridging evolutionary, comparative and neuroethological approaches. Eur. J. Neurosci. 34, 884–907.
|
Basch, M.L., Brown, R.M., Jen, H.I., Groves, A.K., 2016. Where hearing starts: the development of the mammalian cochlea. J. Anat. 228, 233–254.
|
Beatini, J.R., Proudfoot, G.A., Gall, M.D., 2018. Frequency sensitivity in Northern saw-whet owls ( Aegolius acadicus). J. Comp. Physiol. A. 204, 145–154.
|
Beecher, M.D., Brenowitz, E.A., 2005. Functional aspects of song learning in songbirds. Trends Ecol. Evol. 20, 143–149.
|
|
Beuter, K.J., Weiss, R., 1986. Properties of the auditory system in birds and the effectiveness of acoustic scaring signals. In: Proceedings of the International Bird Strike Committee, Copenhagen, Denmark, pp. 60–73.
|
Bolhuis, J.J., Okanoya, K., Scharff, C., 2010. Twitter evolution: converging mechanisms in birdsong and human speech. Nat. Rev. Neurosci. 11, 747–759.
|
Bottjer, S.W., Miesner, E.A., Arnold, A.P., 1984. Forebrain lesions disrupt development but not maintenance of song in passerine birds. Science 224, 901–903.
|
Boumans, T., Vignal, C., Smolders, A., Sijbers, J., Verhoye, M., Van Audekerke, J., et al., 2008. Functional magnetic resonance imaging in zebra finch discerns the neural substrate involved in segregation of conspecific song from background noise. J. Neurophysiol. 99, 931–938.
|
Branch, C.L., Pravosudov, V.V., 2015. Mountain chickadees from different elevations sing different songs: acoustic adaptation, temporal drift or signal of local adaptation? R. Soc. Open. Sci. 2, 150019.
|
|
Brand, A.R., Kellogg, P.P., 1939. Auditory responses of starlings, English sparrows, and domestic pigeons. Wilson Bull. 38–41.
|
Brenowitz, E.A., 2013. Testosterone and brain-derived neurotrophic factor interactions in the avian song control system. Neuroscience 239, 115–123.
|
Brenowitz, E.A., Beecher, M.D., 2005. Song learning in birds: diversity and plasticity, opportunities and challenges. Trends Neurosci. 28, 127–132.
|
Brenowitz, E.A., Larson, T.A., 2015. Neurogenesis in the adult avian song-control system. Cold. Spring. Harb. Perspect. Biol. 7, a019000.
|
Brumm, H., Slater, P.J., 2006. Ambient noise, motor fatigue, and serial redundancy in chaffinch song. Behav. Ecol. Sociobiol. 60, 475–481.
|
Cahill, J.A., Armstrong, J., Deran, A., Khoury, C.J., Paten, B., Haussler, D., et al., 2021. Positive selection in noncoding genomic regions of vocal learning birds is associated with genes implicated in vocal learning and speech functions in humans. Genome Res. 31, 2035–2049.
|
Canopoli, A., Herbst, J.A., Hahnloser, R.H., 2014. A higher sensory brain region is involved in reversing reinforcement-induced vocal changes in a songbird. J. Neurosci. 34, 7018–7026.
|
Caras, M.L., O'Brien, M., Brenowitz, E.A., Rubel, E.W., 2012. Estradiol selectively enhances auditory function in avian forebrain neurons. J. Neurosci. 32, 17597–17611.
|
|
Catchpole, C.K., Slater, P.J., 2003. Bird Song: Biological Themes and Variations. Cambridge University Press, Cambridge.
|
Chang, E.F., Raygor, K.P., Berger, M.S., 2015. Contemporary model of language organization: an overview for neurosurgeons. J. Neurosurg. 122, 250–261.
|
Chen, Z., Ye, R., Goldman, S.A., 2013. Testosterone modulation of angiogenesis and neurogenesis in the adult songbird brain. Neuroscience 239, 139–148.
|
Chung, J.H., Bottjer, S.W., 2022. Developmentally regulated pathways for motor skill learning in songbirds. J. Comp. Neurol. 530, 1288–1301.
|
|
Cohen, S.M., Stebbins, W.C., Moody, D.B., 1978. Audibility thresholds of the blue jay. Auk 95, 563–568.
|
Coleman, M.J., Mooney, R., 2004. Synaptic transformations underlying highly selective auditory representations of learned birdsong. J. Neurosci. 24, 7251–7265.
|
Colquitt, B.M., Merullo, D.P., Konopka, G., Roberts, T.F., Brainard, M.S., 2021. Cellular transcriptomics reveals evolutionary identities of songbird vocal circuits. Science 371, eabd9704.
|
|
Cornez, G., Shevchouk, O.T., Ghorbanpoor, S., Ball, G.F., Cornil, C.A., Balthazart, J., 2020. Testosterone stimulates perineuronal nets development around parvalbumin cells in the adult canary brain in parallel with song crystallization. Horm. Beyond Behav. 119, 104643.
|
Cotanche, D.A., 1987. Regeneration of hair cell stereociliary bundles in the chick cochlea following severe acoustic trauma. Hear. Res. 30, 181–195.
|
Cotanche, D.A., Dopyera, C.E., 1990. Hair cell and supporting cell response to acoustic trauma in the chick cochlea. Hear. Res. 46, 29–40.
|
Courter, J.R., Perruci, R.J., McGinnis, K.J., Rainieri, J.K., 2020. Black-capped chickadees ( Poecile atricapillus) alter alarm call duration and peak frequency in response to traffic noise. PLoS One 15, e0241035.
|
Danish, H.H., Aronov, D., Fee, M.S., 2017. Rhythmic syllable-related activity in a songbird motor thalamic nucleus necessary for learned vocalizations. PLoS One 12, e0169568.
|
|
de Bournonville, C., McGrath, A., Remage-Healey, L., 2020. Testosterone synthesis in the female songbird brain. Horm. Beyond Behav. 121, 104716.
|
Deoniziak, K., Osiejuk, T.S., 2019. Habitat-related differences in song structure and complexity in a songbird with a large repertoire. BMC Ecol. 19, 1–11.
|
Derryberry, E.P., Danner, R.M., Danner, J.E., Derryberry, G.E., Phillips, J.N., Lipshutz, S.E., et al., 2016. Patterns of song across natural and anthropogenic soundscapes suggest that white-crowned sparrows minimize acoustic masking and maximize signal content. PLoS One 11, e0154456.
|
Derryberry, E.P., Luther, D., 2021. What is known—and not known—about acoustic communication in an urban soundscape. Integr. Comp. Biol. 61, 1783–1794.
|
Derryberry, E.P., Phillips, J.N., Derryberry, G.E., Blum, M.J., Luther, D., 2020. Singing in a silent spring: birds respond to a half-century soundscape reversion during the COVID-19 shutdown. Science 370, 575–579.
|
DeVoogd, T., Nottebohm, F., 1981. Gonadal hormones induce dendritic growth in the adult avian brain. Science 214, 202–204.
|
Ding, L., Perkel, D.J., 2002. Dopamine modulates excitability of spiny neurons in the avian basal ganglia. J. Neurosci. 22, 5210–5218.
|
Ding, L., Perkel, D.J., Farries, M.A., 2003. Presynaptic depression of glutamatergic synaptic transmission by D1-like dopamine receptor activation in the avian basal ganglia. J. Neurosci. 23, 6086–6095.
|
Dingle, C., Halfwerk, W., Slabbekoorn, H., 2008. Habitat‐dependent song divergence at subspecies level in the grey‐breasted wood‐wren. J. Evol. Biol. 21, 1079–1089.
|
|
Dooling, R.J., Lohr, B., Dent, M.L., 2000. Hearing in Birds and Reptiles. Springer, New York.
|
Dooling, R.J., Mulligan, J.A., Miller, J.D., 1971. Auditory sensitivity and song spectrum of the common canary ( Serinus canarius). J. Acoust. Soc. Am. 50, 700–709.
|
Dooling, R.J., Peters, S.S., Searcy, M.H., 1979. Auditory sensitivity and vocalizations of the field sparrow ( Spizella pusilla). Bull. Psychon. Soc. 14, 106–108.
|
Dooling, R.J., Popper, A.N., 2016. Some lessons from the effects of highway noise on birds. Proc. Mtgs. Acoust. 27, 010004.
|
Dooling, R.J., Zoloth, S.R., Baylis, J.R., 1978. Auditory sensitivity, equal loudness, temporal resolving power, and vocalizations in the house finch ( Carpodacus mexicanus). J. Comp. Physiol. Psychol. 92, 867.
|
Döppler, J.F., Bush, A., Amador, A., Goller, F., Mindlin, G.B., 2018. Gating related activity in a syringeal muscle allows the reconstruction of zebra finches songs. Chaos 28, 075517.
|
Duque, F.G., Carruth, L.L., 2022. Vocal communication in hummingbirds. Brain Behav. Evol. 97, 241–252.
|
Duque, F.G., Rodriguez-Saltos, C.A., Uma, S., Nasir, I., Monteros, M.F., Wilczynski, W., et al., 2020. High-frequency hearing in a hummingbird. Sci. Adv. 6, eabb9393.
|
Düring, D.N., Ziegler, A., Thompson, C.K., Ziegler, A., Faber, C., Müller, J., et al., 2013. The songbird syrinx morphome: a three-dimensional, high-resolution, interactive morphological map of the zebra finch vocal organ. BMC Biol. 11, 1–27.
|
Edwards, E.P., 1943. Hearing ranges of four species of birds. Auk 60, 239–241.
|
Eens, M., Pinxten, R., Verheyen, R.F., 1991. Male song as a cue for mate choice in the European starling. Behaviour 116, 210–238.
|
Elemans, C.P., Zaccarelli, R., Herzel, H., 2008. Biomechanics and control of vocalization in a non-songbird. J. R. Soc. Interface 5, 691–703.
|
|
Elie, J.E., Hoffmann, S., Dunning, J.L., Coleman, M.J., Fortune, E.S., Prather, J.F., 2020. From perception to action: the role of auditory input in shaping vocal communication and social behaviors in birds. Brain Behav. Evol. 94, 51–60.
|
Engel, M.S., Young, R.J., Davies, W.J., Waddington, D., Wood, M.D., 2024. A systematic review of anthropogenic noise impact on avian species. Conserv. Biol. 10, 684–709.
|
|
Faiß, M., Riede, T., Goller, F., 2022. Tonality over a broad frequency range is linked to vocal learning in birds. Proc. Biol. Sci. 289, 20220792.
|
|
Fernández-Vargas, M., Macedo-Lima, M., Remage-Healey, L., 2024. Acute aromatase inhibition impairs neural and behavioral auditory scene analysis in zebra finches. eNeuro 11, 423.
|
Fischer, B.J., Peña, J.L., 2011. Owl's behavior and neural representation predicted by Bayesian inference. Nat. Neurosci. 14, 1061–1066.
|
|
Fischer, F.P., 1994. General pattern and morphological specializations of the avian cochlea. Scanning Microsc. 8, 18.
|
Fletcher, N.H., 2004. A simple frequency-scaling rule for animal communication. J. Acoust. Soc. Am. 115, 2334–2338.
|
|
Forstmeier, W., Balsby, T.J., 2002. Why mated dusky warblers sing so much: territory guarding and male quality announcement. Behaviour, 89–111.
|
Fortune, E.S., Rodríguez, C., Li, D., Ball, G.F., Coleman, M.J., 2011. Neural mechanisms for the coordination of duet singing in wrens. Science 334, 666–670.
|
Friis, J.I., Sabino, J., Santos, P., Dabelsteen, T., Cardoso, G.C., 2021. The allometry of sound frequency bandwidth in songbirds. Am. Nat. 197, 607–614.
|
Frohlich, F., Basta, D., Strübing, I., Ernst, A., Gröschel, M., 2017. Time course of cell death due to acoustic overstimulation in the mouse medial geniculate body and primary auditory cortex. Noise Health 19, 133–139.
|
Gale, S.D., Perkel, D.J., 2006. Physiological properties of zebra finch ventral tegmental area and substantia nigra pars compacta neurons. J. Neurosci. 96, 2295–2306.
|
Gentner, T.Q., Hulse, S.H., Bentley, G.E., Ball, G.F., 2000. Individual vocal recognition and the effect of partial lesions to HVc on discrimination, learning, and categorization of conspecific song in adult songbirds. J. Neurobiol. 42, 117–133.
|
Gentry, K.E., Derryberry, E.P., Danner, R.M., Danner, J.E., Luther, D.A., 2017. Immediate signaling flexibility in response to experimental noise in urban, but not rural, white‐crowned sparrows. Ecosphere 8, e01916.
|
Gentry, K.E., Luther, D.A., 2019. Noise-induced vocal plasticity in urban white-crowned sparrows does not involve adjustment of trill performance components. Sci. Rep. 9, 1905.
|
Gobes, S.M., Bolhuis, J.J., 2007. Birdsong memory: a neural dissociation between song recognition and production. Curr. Biol. 17, 789–793.
|
Goller, F., 2022. The syrinx. Curr. Biol. 32, R1095–R1100.
|
Goller, F., Riede, T., 2013. Integrative physiology of fundamental frequency control in birds. J. Physiol. Paris 107, 230–242.
|
Goller, F., Suthers, R.A., 1996. Role of syringeal muscles in controlling the phonology of bird song. J. Neurophysiol. 76, 287–300.
|
Gough, D.C., Mennill, D.J., Nol, E., 2014. Singing seaside: pacific Wrens ( Troglodytes pacificus) change their songs in the presence of natural and anthropogenic noise. Wilson J. Ornithol. 126, 269–278.
|
Grimes, S.E., Lewis, E.J., Nduwimana, L.A., Yurk, B., Ronald, K.L., 2024. Urbanization alters the song propagation of two human-commensal songbird species. J. Acoust. Soc. Am. 155, 2803–2816.
|
|
Grothe, B., Carr, C.E., Casseday, J.H., Fritzsch, B., Köppl, C., 2004. The evolution of central pathways and their neural processing patterns. Evolution of the Vertebrate Auditory System, pp. 289–359.
|
Guo, F., Bonebrake, T.C., Dingle, C., 2016. Low frequency dove coos vary across noise gradients in an urbanized environment. Behav. Process. 129, 86–93.
|
Hahnloser, R.H., Kozhevnikov, A.A., Fee, M.S., 2002. An ultra-sparse code underliesthe generation of neural sequences in a songbird. Nature 419, 65–70.
|
Harding, C.F., 2004. Hormonal modulation of singing: hormonal modulation of the songbird brain and singing behavior. Ann. N. Y. Acad. Sci. 1016, 524–539.
|
Hardt, B., Benedict, L., 2021. Can you hear me now? A review of signal transmission and experimental evidence for the acoustic adaptation hypothesis. Bioacoustics 30, 716–742.
|
|
Hay, E.M., McGee, M.D., White, C.R., Chown, S.L., 2024. Body size shapes song in honeyeaters. Proc. Biol. Sci. 291, 20240339.
|
Heberden, C., 2017. Sex steroids and neurogenesis. Biochem. Pharmacol. 141, 56–62.
|
Heffner, R., Cumming, J.F., Koay, G., Heffner, H.E., 2020. Hearing in Indian peafowl ( Pavo cristatus): sensitivity to infrasound. J. Comp. Physiol. A. 206, 899–906.
|
Heffner, H.E., Koay, G., Heffner, R.S., 2016. Budgerigars ( Melopsittacus undulatus) do not hear infrasound: the audiogram from 8 Hz to 10 kHz. J. Comp. Physiol. A. 202, 853–857.
|
Heffner, H.E., Koay, G., Heffner, R.S., 2024. Hearing in helmeted guineafowl ( Numida meleagris): audiogram from 2 Hz to 10 kHz and localization acuity for brief noise bursts. J. Comp. Physiol. A. 210, 65–73.
|
Heffner, H.E., Koay, G., Hill, E.M., Heffner, R.S., 2013. Conditioned suppression/avoidance as a procedure for testing hearing in birds: the domestic pigeon ( Columba livia). Behav. Res. Methods 45, 383–392.
|
|
Henry, K.S., Gall, M.D., Vélez, A., Lucas, J.R., 2016. Avian auditory processing at four different scales: variation among species, seasons, sexes, and individuals. Psychological Mechanisms in Animal Communication, pp. 17–55.
|
Hienz, R.D., Sinnott, J.M., Sachs, M.B., 1977. Auditory sensitivity of the redwing Blackbird (Agelaius phoeniceus) and brown-headed cowbird ( Molothrus ater). J. Comp. Physiol. Psychol. 91, 1365.
|
Hill, E.M., 2017. Audiogram of the mallard duck ( Anas platyrhynchos) from 16 Hz to 9 kHz. J. Comp. Physiol. A. 203, 929–934.
|
Hill, S.D., Aryal, A., Pawley, M.D., Ji, W., 2018. So much for the city: urban–rural song variation in a widespread Asiatic songbird. Integr. Zool. 13, 194–205.
|
Hill, E.M., Koay, G., Heffner, R.S., Heffner, H.E., 2014. Audiogram of the chicken ( Gallus gallus domesticus) from 2 Hz to 9 kHz. J. Comp. Physiol. A. 200, 863–870.
|
Hirokawa, N., 1978. The ultrastructure of the basilar papilla of the chick. J. Comp. Neurol. 181, 361–374.
|
Hoffmann, L.A., Saravanan, V., Wood, A.N., He, L., Sober, S.J., 2016. Dopaminergic contributions to vocal learning. J. Neurosci. 36, 2176–2189.
|
Hudspeth, A.J., Choe, Y., Mehta, A.D., Martin, P., 2000. Putting ion channels to work: mechanoelectrical transduction, adaptation, and amplification by hair cells. Proc. Natl. Acad. Sci. U. S. A. 97, 11765–11772.
|
Husbands, J.M., Steinberg, S.A., Kurian, R., Saunders, J.C., 1999. Tip-link integrity on chick tall hair cell stereocilia following intense sound exposure. Hear. Res. 135, 135–145.
|
Hynes, D.P., Miller, E.H., 2014. Vocal distinctiveness of the red crossbill ( Loxia curvirostra) on the island of newfoundland, Canada. Auk. 131, 421–433.
|
Inda, M., Hotta, K., Oka, K., 2020. Neural properties of fundamental function encoding of sound selectivity in the female avian auditory cortex. Eur. J. Neurosci. 51, 1770–1783.
|
Ivanitskii, V.V., Marova, I.M., 2022. The syntactic organization of bird song. Biol. Bull. 49, 1158–1170.
|
Janes, S.W., Ryker, L., 2013. Rapid change in a type I song dialect of Hermit Warblers ( Setophaga occidentalis). Auk 130, 30–35.
|
|
Janesick, A., Scheibinger, M., Benkafadar, N., Kirti, S., Ellwanger, D.C., Heller, S., 2021. Cell-type identity of the avian cochlea. Cell Rep. 34.
|
Jarvis, E.D., Ribeiro, S., Da Silva, M.L., Ventura, D., Vielliard, J., Mello, C.V., 2000. Behaviourally driven gene expression reveals song nuclei in hummingbird brain. Nature 406, 628–632.
|
Jeanne, J.M., Thompson, J.V., Sharpee, T.O., Gentner, T.Q., 2011. Emergence of learned categorical representations within an auditory forebrain circuit. J. Neurosci. 31, 2595–2606.
|
Jeong, J.K., Burrows, K., Tremere, L.A., Pinaud, R., 2011. Neurochemical organization and experience‐dependent activation of estrogen‐associated circuits in the songbird auditory forebrain. Eur. J. Neurosci. 34, 283–291.
|
|
Job, J.R., Kohler, S.L., Gill, S.A., 2016. Song adjustments by an open habitat bird to anthropogenic noise, urban structure, and vegetation. Behav. Ecol. 27, 1734–1744.
|
|
Kang, H., Dos Santos, E.B., Kojima, S., 2025. Neural sensitivity to frequency changes in song structure in a high-order auditory area reflects tutor song memory in adult songbirds. Brain Struct. Funct. 230, 1–9.
|
Kao, M.H., Brainard, M.S., 2006. Lesions of an avian basal ganglia circuit prevent context-dependent changes to song variability. J. Neurophysiol. 96, 1441–1455.
|
Kao, M.H., Wright, B.D., Doupe, A.J., 2008. Neurons in a forebrain nucleus required for vocal plasticity rapidly switch between precise firing and variable bursting depending on social context. J. Neurosci. 28, 13232–13247.
|
Katic, J., Morohashi, Y., Yazaki-Sugiyama, Y., 2022. Neural circuit for social authentication in song learning. Nat. Commun. 13, 4442.
|
Keen, S.C., Cole, E.F., Sheehan, M.J., Sheldon, B.C., 2020. Social learning of acoustic anti-predator cues occurs between wild bird species. Proc. Biol. Sci. 287, 20192513.
|
Kight, C.R., Swaddle, J.P., 2015. Eastern bluebirds alter their song in response to anthropogenic changes in the acoustic environment. Integr. Comp. Biol. 55, 418–431.
|
Kingsley, E.P., Eliason, C.M., Riede, T., Li, Z., Hiscock, T.W., Farnsworth, M., et al., 2018. Identity and novelty in the avian syrinx. Proc. Natl. Acad. Sci. U. S. A. 115, 10209–10217.
|
Kraus, K.S., Mitra, S., Jimenez, Z., Hinduja, S., Ding, D., Jiang, H., et al., 2010. Noise trauma impairs neurogenesis in the rat hippocampus. Neuroscience 167, 1216–1226.
|
Kroodsma, D.E., Houlihan, P.W., Falleon, P.A., Wells, J.A., 1997. Song development by grey catbirds. Anim. Behav. 54, 457–464.
|
Krumm, B., Klump, G., Köppl, C., Langemann, U., 2017. Barn owls have ageless ears. Proc. Biol. Sci. 284, 20171584.
|
Kuebrich, B.D., Sober, S.J., 2015. Variations on a theme: songbirds, variability, and sensorimotor error correction. Neuroscience 296, 48–54.
|
Kunc, H.P., Morrison, K., Schmidt, R., 2022. A meta-analysis on the evolution of the Lombard effect reveals that amplitude adjustments are a widespread vertebrate mechanism. Proc. Natl. Acad. Sci. U. S. A. 119, e2117809119.
|
Larson, T.A., Thatra, N.M., Lee, B.H., Brenowitz, E.A., 2014. Reactive neurogenesis in response to naturally occurring apoptosis in an adult brain. J. Neurosci. 34, 13066–13076.
|
Larson, T.A., Wang, T.W., Gale, S.D., Miller, K.E., Thatra, N.M., Caras, M.L., et al., 2013. Postsynaptic neural activity regulates neuronal addition in the adult avian song control system. Proc. Natl. Acad. Sci. U. S. A. 110, 16640–16644.
|
|
Las, L., Fee, M., 2008. Recordings of striatal-projecting neurons in the ventral tegmental area (VTA) of the juvenile zebra finch during song learning. In: Society for Neuroscience Annual Meeting Abstracts.
|
Lawley, K.S., Fenn, T., Person, E., Huber, H., Zaharas, K., Smith, P., et al., 2022. Auditory processing neurons influence song evaluation and strength of mate preference in female songbirds. Front. Neural Circ. 16, 994548.
|
Leblois, A., Wendel, B.J., Perkel, D.J., 2010. Striatal dopamine modulates basal ganglia output and regulates social context-dependent behavioral variability through D1 receptors. J. Neurosci. 30, 5730–5743.
|
Lengagne, T., Aubin, T., Lauga, J., Jouventin, P., 1999. How do king penguins ( Aptenodytes patagonicus) apply the mathematical theory of information to communicate in windy conditions? Proc. Roy. Soc. Lond. B. 266, 1623–1628.
|
Lengagne, T., Slater, P.J., 2002. The effects of rain on acoustic communication: tawny owls have good reason for calling less in wet weather. Proc. Roy. Soc. Lond. B. 269, 2121–2125.
|
Lewandowski, B., Vyssotski, A., Hahnloser, R.H., Schmidt, M., 2013. At the interface of the auditory and vocal motor systems: NIf and its role in vocal processing, production and learning. J. Physiol. Paris 107, 178–192.
|
Li, X.C., Jarvis, E.D., Alvarez-Borda, B., Lim, D.A., Nottebohm, F., 2000. A relationship between behavior, neurotrophin expression, and new neuron survival. Proc. Natl. Acad. Sci. U. S. A. 97, 8584–8589.
|
Liang, B., Li, Y., Zhao, W., Du, Y., 2023. Bilateral human laryngeal motor cortex in perceptual decision of lexical tone and voicing of consonant. Nat. Commun. 14, 4710.
|
Lipkind, D., Nottebohm, F., Rado, R., Barnea, A., 2002. Social change affects the survival of new neurons in the forebrain of adult songbirds. Behav. Brain Res. 133, 31–43.
|
Louder, M.I., Lawson, S., Lynch, K.S., Balakrishnan, C.N., Hauber, M.E., 2019. Neural mechanisms of auditory species recognition in birds. Biol. Rev. 94, 1619–1635.
|
Lucas, J., Freeberg, T., Krishnan, A., Long, G., 2002. A comparative study of avian auditory brainstem responses: correlations with phylogeny and vocal complexity, and seasonal effects. J. Comp. Physiol. A. 188, 981–992.
|
Luther, D.A., Derryberry, E.P., 2012. Birdsongs keep pace with city life: changes in song over time in an urban songbird affects communication. Anim. Behav. 83, 1059–1066.
|
Lynch, K.S., Louder, M.I., Hauber, M.E., 2018. Species-specific auditory forebrain responses to non-learned vocalizations in juvenile blackbirds. Brain Behav. Evol. 91, 193–200.
|
Lynch, G.F., Okubo, T.S., Hanuschkin, A., Hahnloser, R.H., Fee, M.S., 2016. Rhythmic continuous-time coding in the songbird analog of vocal motor cortex. Neuron 90, 877–892.
|
Mack, A.L., Jones, J., 2003. Low-frequency vocalizations by cassowaries ( Casuarius spp.). Auk 120, 1062–1068.
|
Mann, D.C., Fitch, W.T., Tu, H.W., Hoeschele, M., 2021. Universal principles underlying segmental structures in parrot song and human speech. Sci. Rep. 11, 1–13.
|
Maldarelli, G., Firzlaff, U., Kettler, L., Ondracek, J.M., Luksch, H., 2022. Two types of auditory spatial receptive fields in different parts of the chicken's midbrain. J. Neurosci. 42, 4669–4680.
|
|
Manohar, S., Chen, G.D., Ding, D., Liu, L., Wang, J., Chen, Y.C., et al., 2022. Unexpected consequences of noise-induced hearing loss: impaired hippocampal neurogenesis, memory, and stress. Front. Integr. Neurosci 16, 871223.
|
Manukyan, A.L., 2022. Noise as a cause of neurodegenerative disorders: molecular and cellular mechanisms. Neurol. Sci. 43, 2983–2993.
|
Marean, G.C., Burt, J.M., Beecher, M.D., Rubel, E.W., 1993. Hair cell regeneration in the European starling ( Sturnus vulgaris): recovery of pure-tone detection thresholds. Hear. Res. 71, 125–136.
|
|
Marler, P.R., Slabbekoorn, H., 2004. Nature’s Music: the Science of Birdsong. Elsevier.
|
Marten, K., Quine, D., Marler, P., 1977. Sound transmission and its significance for animal vocalization: Ⅱ. Tropical forest habitats. Behav. Ecol. Sociobiol. 2, 291–302.
|
McMullen, H., Schmidt, R., Kunc, H.P., 2014. Anthropogenic noise affects vocal interactions. Behav. Process. 103, 125–128.
|
McPherson, D.R., 2018. Sensory hair cells: an introduction to structure and physiology. Integr. Comp. Biol. 58, 282–300.
|
Mello, C.V., Velho, T.A., Pinaud, R., 2004. Song‐induced gene expression: a window on song auditory processing and perception. Ann. N. Y. Acad. Sci. 1016, 263–281.
|
Mikula, P., Valcu, M., Brumm, H., Bulla, M., Forstmeier, W., Petrusková, T., et al., 2021. A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecol. Lett. 24, 477–486.
|
Miller, M.N., Cheung, C.Y.J., Brainard, M.S., 2017. Vocal learning promotes patterned inhibitory connectivity. Nat. Commun. 8, 2105.
|
Mooney, R., 2009. Neurobiology of song learning. Curr. Opin. Neurobiol. 19, 654–660.
|
Mooney, R., Prather, J.F., 2005. The HVC microcircuit: the synaptic basis for interactions between song motor and vocal plasticity pathways. J. Neurosci. 25, 1952–1964.
|
Mooney, T.A., Smith, A., Hansen, K.A., Larsen, O.N., Wahlberg, M., Rasmussen, M., 2019. Birds of a feather: hearing and potential noise impacts in puffins ( Fratercula arctica). Proc. Mtgs. Acoust. 37, 010004.
|
Moore, R.C., Lee, T., Theunissen, F.E., 2013. Noise-invariant neurons in the avian auditory cortex: hearing the song in noise. PLoS Comput. Biol. 9, e1002942.
|
Morton, E.S., 1975. Ecological sources of selection on avian sounds. Am. Nat. 109, 17–34.
|
Moseley, D.L., Derryberry, G.E., Phillips, J.N., Danner, J.E., Danner, R.M., Luther, D.A., et al., 2018. Acoustic adaptation to city noise through vocal learning by a songbird. Proc. Biol. Sci. 285, 20181356.
|
|
Mountjoy, D.J., Lemon, R.E., 1991. Song as an attractant for male and female European starlings, and the influence of song complexity on their response. Behav. Ecol. Sociobiol. 28, 97–100.
|
Nemeth, E., Pieretti, N., Zollinger, S.A., Geberzahn, N., Partecke, J., Miranda, A.C., et al., 2013. Bird song and anthropogenic noise: vocal constraints may explain why birds sing higher-frequency songs in cities. Proc. R. Soc. B. Biol. Sci. 280, 20122798.
|
Nieder, A., Mooney, R., 2020. The neurobiology of innate, volitional and learned vocalizations in mammals and birds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375, 20190054.
|
Nishikawa, J., Okada, M., Okanoya, K., 2008. Population coding of song element sequence in the Bengalese finch HVC. Eur. J. Neurosci. 27, 3273–3283.
|
Nordt, A., Klenke, R., 2013. Sleepless in town–drivers of the temporal shift in dawn song in urban European blackbirds. PLoS One 8, e71476.
|
|
Nowicki, S., 1997. Bird acoustics. In: Crocker, M.J. (Ed.), Encyclopedia of Acoustics. John Wiley Sons, pp. 1813–1817.
|
Nowicki, S., Searcy, W.A., 2004. Song function and the evolution of female preferences: why birds sing, why brains matter. Ann. N. Y. Acad. Sci. 1016, 704–723.
|
|
Peng, A.W., Ricci, A.J., 2011. Somatic motility and hair bundle mechanics, are both necessary for cochlear amplification? Hear. Res. 273, 109–122.
|
Pfenning, A.R., Hara, E., Whitney, O., Rivas, M.V., Wang, R., Roulhac, P.L., et al., 2014. Convergent transcriptional specializations in the brains of humans and song-learning birds. Science 346, 1256846.
|
Phillips, J.N., Derryberry, E.P., 2017. Vocal performance is a salient signal for male–male competition in White-crowned Sparrows. Auk 134, 564–574.
|
Pinaud, R., Terleph, T.A., 2008. A songbird forebrain area potentially involved in auditory discrimination and memory formation. J. Biosci. 33, 145–155.
|
Podos, J., Huber, S.K., Taft, B., 2004. Bird song: the interface of evolution and mechanism. Annu. Rev. Ecol. Evol. Syst. 35, 55–87.
|
Prather, J.F., 2013. Auditory signal processing in communication: perception and performance of vocal sounds. Hear. Res. 305, 144–155.
|
Prather, J.F., Peters, S., Nowicki, S., Mooney, R., 2008. Precise auditory–vocal mirroring in neurons for learned vocal communication. Nature 451, 305–310.
|
Pytte, C.L., George, S., Korman, S., David, E., Bogdan, D., Kirn, J.R., 2012. Adult neurogenesis is associated with the maintenance of a stereotyped, learned motor behavior. J. Neurosci. 32, 7052–7057.
|
Pytte, C.L., Parent, C., Wildstein, S., Varghese, C., Oberlander, S., 2010. Deafening decreases neuronal incorporation in the zebra finch caudomedial nidopallium (NCM). Behav. Brain Res. 211, 141–147.
|
Pytte, C., Yu, Y.L., Wildstein, S., George, S., Kirn, J.R., 2011. Adult neuron addition to the zebra finch song motor pathway correlates with the rate and extent of recovery from botox-induced paralysis of the vocal muscles. J. Neurosci. 31, 16958–16968.
|
Reed, V.A., Toth, C.A., Wardle, R.N., Gomes, D.G., Barber, J.R., Francis, C.D., 2022. Experimentally broadcast ocean surf and river noise alters birdsong. PeerJ 10, e13297.
|
Remage-Healey, L., Maidment, N.T., Schlinger, B.A., 2008. Forebrain steroid levels fluctuate rapidly during social interactions. Nat. Neurosci. 11, 1327–1334.
|
Riede, T., Suthers, R.A., Fletcher, N.H., Blevins, W.E., 2006. Songbirds tune their vocal tract to the fundamental frequency of their song. Proc. Natl. Acad. Sci. U. S. A. 103, 5543–5548.
|
Roberts, T.F., Mooney, R., 2013. Motor circuits help encode auditory memories of vocal models used to guide vocal learning. Hear. Res. 303, 48–57.
|
Rose, E.M., Prior, N.H., Ball, G.F., 2022. The singing question: Re‐conceptualizing birdsong. Biol. Rev. Camb. Phil. Soc. 97, 326–342.
|
Rosen, M.J., Mooney, R., 2006. Synaptic interactions underlying song-selectivity in the avian nucleus HVC revealed by dual intracellular recordings. J. Neurophysiol. 95, 1158–1175.
|
Rubel, E.W., Furrer, S.A., Stone, J.S., 2013. A brief history of hair cell regeneration research and speculations on the future. Hear. Res. 297, 42–51.
|
Sainburg, T., Theilman, B., Thielk, M., Gentner, T.Q., 2019. Parallels in the sequential organization of birdsong and human speech. Nat. Commun. 10, 3636.
|
Sanford, S.E., Lange, H.S., Maney, D.L., 2010. Topography of estradiol‐modulated genomic responses in the songbird auditory forebrain. Dev. Neurobiol. 70, 73–86.
|
|
Saravanan, V., Hoffmann, L.A., Jacob, A.L., Berman, G.J., Sober, S.J., 2019. Dopamine depletion affects vocal acoustics and disrupts sensorimotor adaptation in songbirds. eNeuro 6, 190.
|
Sasaki, A., Sotnikova, T.D., Gainetdinov, R.R., Jarvis, E.D., 2006. Social context-dependent singing-regulated dopamine. J. Neurosci. 26, 9010–9014.
|
Sato, M.P., Benkafadar, N., Heller, S., 2024. Hair cell regeneration, reinnervation, and restoration of hearing thresholds in the avian hearing organ. Cell Rep. 43, 113822.
|
Schmidt, M.F., 2003. Pattern of interhemispheric synchronization in HVc during singing correlates with key transitions in the song pattern. J. Neurophysiol. 90, 3931–3949.
|
Schmidt, M.F., McLean, J., Goller, F., 2012. Breathing and vocal control: the respiratory system as both a driver and a target of telencephalic vocal motor circuits in songbirds. Exp. Physiol. 97, 455–461.
|
|
Schneider, D.M., Woolley, S.M., 2009. Stimulus-dependent receptive field dynamics are driven by subthreshold excitation and natural stimulus statistics. Society for Neuroscience Annual Meeting Abstracts.
|
Schroeder, K.M., Remage‐Healey, L., 2021. Adult‐like neural representation of species‐specific songs in the auditory forebrain of zebra finch nestlings. Dev. Neurobiol. 81, 123–138.
|
|
Schumacher, J.W., Woolley, S.M., 2009. Encoding properties of midbrain neurons in awake and anesthetized songbirds. Society for Neuroscience Annual Meeting Abstracts.
|
Scotto‐Lomassese, S., Rochefort, C., Nshdejan, A., Scharff, C., 2007. HVC interneurons are not renewed in adult male zebra finches. Eur. J. Neurosci. 25, 1663–1668.
|
Scully, E.N., Hahn, A.H., Campbell, K.A., McMillan, N., Congdon, J.V., Sturdy, C.B., 2017. ZENK expression following conspecific and heterospecific playback in the zebra finch auditory forebrain. Behav. Brain Res. 331, 151–158.
|
Sebastianelli, M., Blumstein, D.T., Kirschel, A.N., 2020. Ambient noise from ocean surf drives frequency shifts in non-passerine bird song. bioRxiv. .
|
Sekiya, T., Viberg, A., Kojima, K., Sakamoto, T., Nakagawa, T., Ito, J., et al., 2012. Trauma‐specific insults to the cochlear nucleus in the rat. J. Neurosci. Res. 90, 1924–1931.
|
Shaevitz, S.S., Theunissen, F.E., 2007. Functional connectivity between auditory areas field L and CLM and song system nucleus HVC in anesthetized zebra finches. J. Neurophysiol. 98, 2747–2764.
|
Shevchouk, O.T., Ball, G.F., Cornil, C.A., Balthazart, J., 2017. Studies of HVC plasticity in adult canaries reveal social effects and sex differences as well as limitations of multiple markers available to assess adult neurogenesis. PLoS One 12, e0170938.
|
Shizuka, D., Lein, M.R., Chilton, G., 2016. Range-wide patterns of geographic variation in songs of Golden-crowned Sparrows ( Zonotrichia atricapilla). Auk 133, 520–529.
|
Simmonds, A.J., Leech, R., Iverson, P., Wise, R.J., 2014. The response of the anterior striatum during adult human vocal learning. J. Neurophysiol. 112, 792–801.
|
Sisneros, J.A., Forlano, P.M., Deitcher, D.L., Bass, A.H., 2004. Steroid-dependent auditory plasticity leads to adaptive coupling of sender and receiver. Science 305, 404–407.
|
Slabbekoorn, H., Peet, M., 2003. Birds sing at a higher pitch in urban noise. Nature 424, 267.
|
|
Slabbekoorn, H., Smith, T.B., 2002. Habitat‐dependent song divergence in the little greenbul: an analysis of environmental selection pressures on acoustic signals. Evolution 56, 1849–1858.
|
Smolders, J.W.T., 1999. Functional recovery in the avian ear after hair cell regeneration. Audiol. Neurootol. 4, 286–302.
|
Snell-Rood, E.C., 2012. The effect of climate on acoustic signals: does atmospheric sound absorption matter for bird song and bat echolocation? J. Acoust. Soc. Am. 131, 1650–1658.
|
Solis, M.M., Brainard, M.S., Hessler, N.A., Doupe, A.J., 2000. Song selectivity and sensorimotor signals in vocal learning and production. Proc. Natl. Acad. Sci. U. S. A. 97, 11836–11842.
|
|
Stewart, P.A., 1955. An audibility curve for two ring-necked pheasants. Ohio. J. Sci. 55, 122–125.
|
Strawn, S.N., Hill, E.M., 2020. Japanese quail ( Coturnix japonica) audiogram from 16 Hz to 8 kHz. J. Comp. Physiol. A. 206, 665–670.
|
Tachibana, R.O., Lee, D., Kai, K., Kojima, S., 2022. Performance-dependent consolidation of learned vocal changes in adult songbirds. J. Neurosci. 42, 1974–1986.
|
Takahashi, T.T., 2010. How the owl tracks its prey – Ⅱ. J. Exp. Biol. 213, 3399–3408.
|
Tang, Y., Christensen‐Dalsgaard, J., Carr, C.E., 2012. Organization of the auditory brainstem in a lizard, Gekko gecko. I. Auditory nerve, cochlear nuclei, and superior olivary nuclei. J. Comp. Neurol. 520, 1784–1799.
|
Tecumseh Fitch, W., Reby, D., 2001. The descended larynx is not uniquely human. Proc. Biol. Sci. 268, 1669–1675.
|
Terpstra, N.J., Bolhuis, J.J., den Boer-Visser, A.M., 2004. An analysis of the neural representation of birdsong memory. J. Neurosci. 24, 4971–4977.
|
Theunissen, F.E., Amin, N., Shaevitz, S.S., Woolley, S.M., Fremouw, T., Hauber, M.E., 2004. Song selectivity in the song system and in the auditory forebrain. Ann. N. Y. Acad. Sci. 1016, 222–245.
|
Thompson, C.K., Meitzen, J., Replogle, K., Drnevich, J., Lent, K.L., Wissman, A.M., et al., 2012. Seasonal changes in patterns of gene expression in avian song control brain regions. PLoS One 7, e35119.
|
Tremere, L.A., Pinaud, R., 2011. Brain-generated estradiol drives long-term optimization of auditory coding to enhance the discrimination of communication signals. J. Neurosci. 31, 3271–3289.
|
Tschida, K., Mooney, R., 2012. The role of auditory feedback in vocal learning and maintenance. Curr. Opin. Neurobiol. 22, 320–327.
|
Vates, G.E., Broome, B.M., Mello, C.V., Nottebohm, F., 1996. Auditory pathways of caudal telencephalon and their relation to the song system of adult male zebra finches ( Taenopygia guttata). J. Comp. Neurol. 366, 613–642.
|
Vernes, S.C., Janik, V.M., Fitch, W.T., Slater, P.J., 2021. Vocal learning in animals and humans. Trans. R. Soc. B. 376, 20200234.
|
|
Voigt, C., Leitner, S., Gahr, M., 2006. Repertoire and structure of duet and solo songs in cooperatively breeding white-browed sparrow weavers. Behaviour, 159–182.
|
Wang, N., Hurley, P., Pytte, C., Kirn, J.R., 2002. Vocal control neuron incorporation decreases with age in the adult zebra finch. J. Neurosci. 22, 10864–10870.
|
Warchol, M.E., 2011. Sensory regeneration in the vertebrate inner ear: differences at the levels of cells and species. Hear. Res. 273, 72–79.
|
Warren, T.L., Tumer, E.C., Charlesworth, J.D., Brainard, M.S., 2011. Mechanisms and time course of vocal learning and consolidation in the adult songbird. J. Neurophysiol. 106, 1806–1821.
|
|
Webster, M.S., Podos, J., 2018. Acoustic communication. In: Morrison, M.L., Rodewald, A.D., Voelker, G., Colon, M.R., Prather, J.F. (Eds.), Ornithology: Foundation, Analysis, and Application. Johns Hopkins University Press, pp. 409–436.
|
Wever, E.G., Herman, P.N., Simmons, J.A., Hertzler, D.R., 1969. Hearing in the blackfooted penguin, Spheniscus demersus, as represented by the cochlear potentials. Proc. Natl. Acad. Sci. U. S. A. 63, 676–680.
|
Wilbrecht, L., Kirn, J.R., 2004. Neuron addition and loss in the song system: regulation and function. Ann. N. Y. Acad. Sci. 1016, 659–683.
|
Wiley, R.H., Richards, D.G., 1978. Physical constraints on acoustic communication in the atmosphere: implications for the evolution of animal vocalizations. Behav. Ecol. Sociobiol. 3, 69–94.
|
Woolley, S.M., Casseday, J.H., 2005. Processing of modulated sounds in the zebra finch auditory midbrain: responses to noise, frequency sweeps, and sinusoidal amplitude modulations. J. Neurophysiol. 94, 1143–1157.
|
Woolley, S.C., Doupe, A.J., 2008. Social context–induced song variation affects female behavior and gene expression. PLoS. Biol 6, e62.
|
Woolley, S.M., Moore, J.M., 2011. Coevolution in communication senders and receivers: vocal behavior and auditory processing in multiple songbird species. Ann. N. Y. Acad. Sci. 1225, 155–165.
|
Woolley, S.M., Wissman, A.M., Rubel, E.W., 2001. Hair cell regeneration and recovery of auditory thresholds following aminoglycoside ototoxicity in Bengalese finches. Hear. Res. 153, 181–195.
|
Xia, A., Liu, X., Raphael, P.D., Applegate, B.E., Oghalai, J.S., 2016. Hair cell force generation does not amplify or tune vibrations within the chicken basilar papilla. Nat. Commun. 7, 13133.
|
|
Yadav, S., Rab, S., Wan, M., Yadav, D., Singh, V.R., 2024. Sound communication in nature. In: Garg, N., Gautam, C., Rab, S., Wan, M., Agarwal, R., Yadav, S. (Eds.), Handbook of Vibroacoustics, Noise and Harshness. Springer Nature Singapore, Singapore, pp. 951–976.
|
Yanagihara, S., Ikebuchi, M., Mori, C., Tachibana, R.O., Okanoya, K., 2021. Neural correlates of vocal initiation in the VTA/SNc of juvenile male zebra finches. Sci. Rep. 11, 22388.
|
Yip, Z.C., Miller‐Sims, V.C., Bottjer, S.W., 2012. Morphology of axonal projections from the high vocal center to vocal motor cortex in songbirds. J. Comp. Neurol. 520, 2742–2756.
|
Yoder, K.M., Vicario, D.S., 2012. To modulate and be modulated: estrogenic influences on auditory processing of communication signals within a socio-neuro-endocrine framework. Behav. Neurosci. 126, 17.
|
Zhang, Y., Zhou, L., Zuo, J., Wang, S., Meng, W., 2023. Analogies of human speech and bird song: from vocal learning behavior to its neural basis. Front. Psychol. 14, 1100969.
|

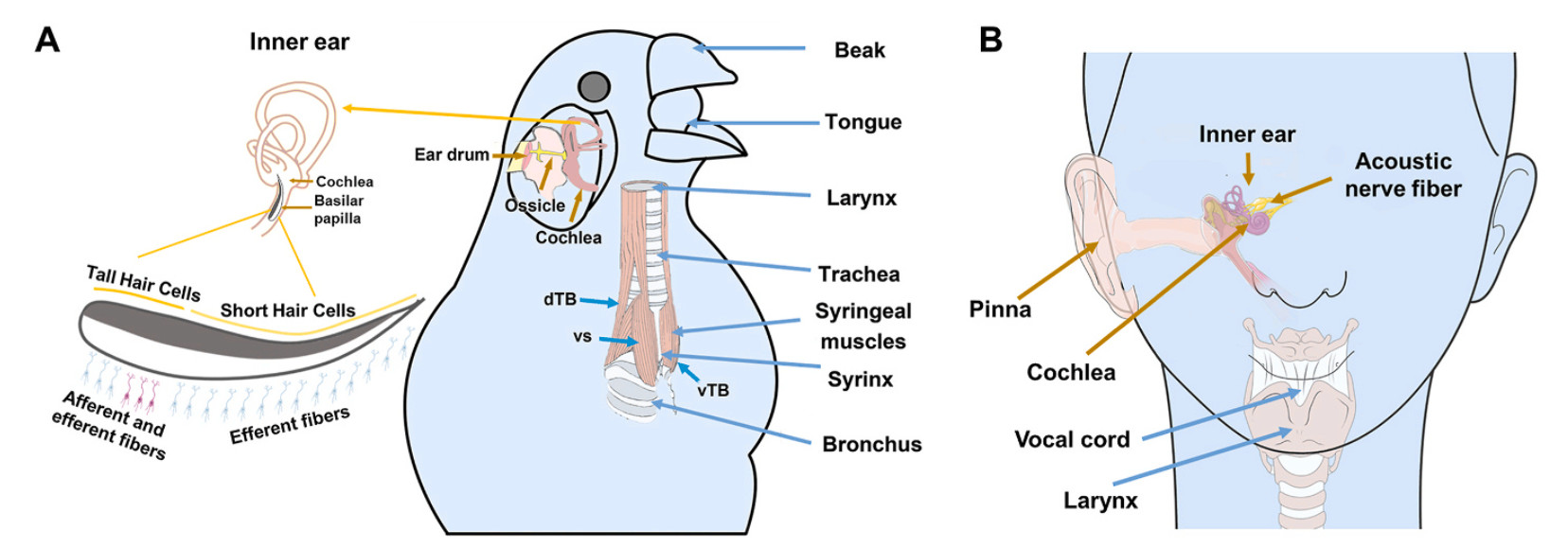
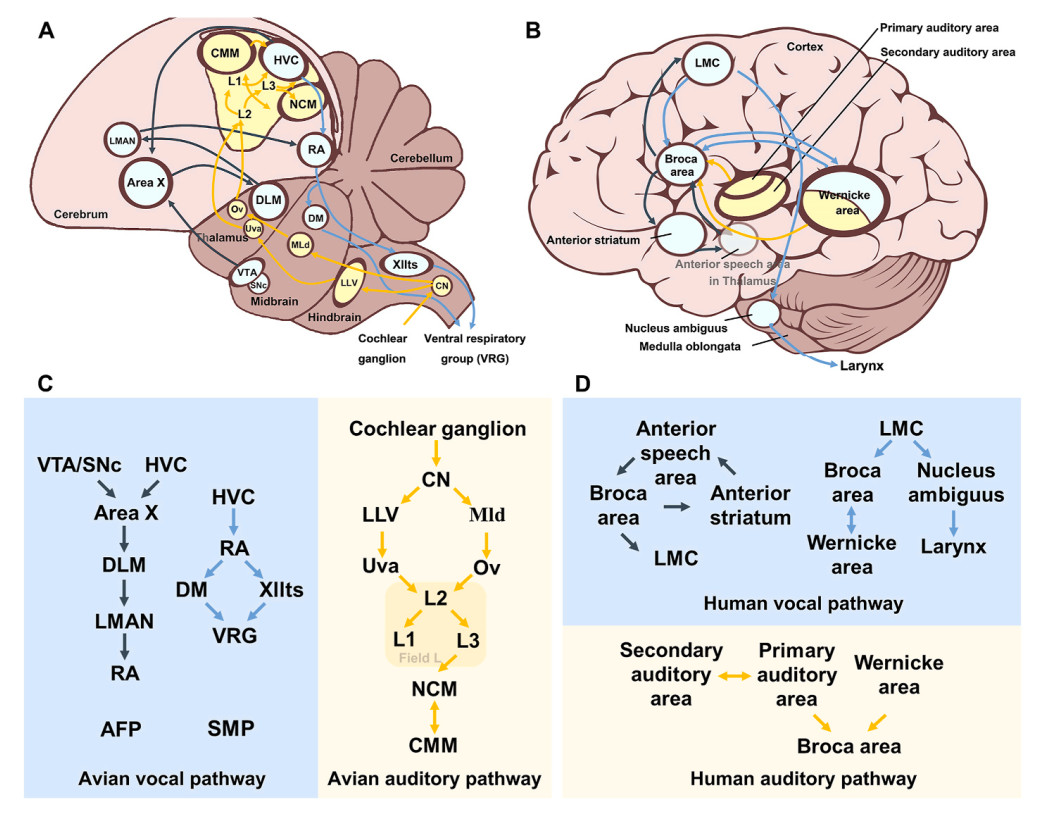
 DownLoad:
DownLoad:
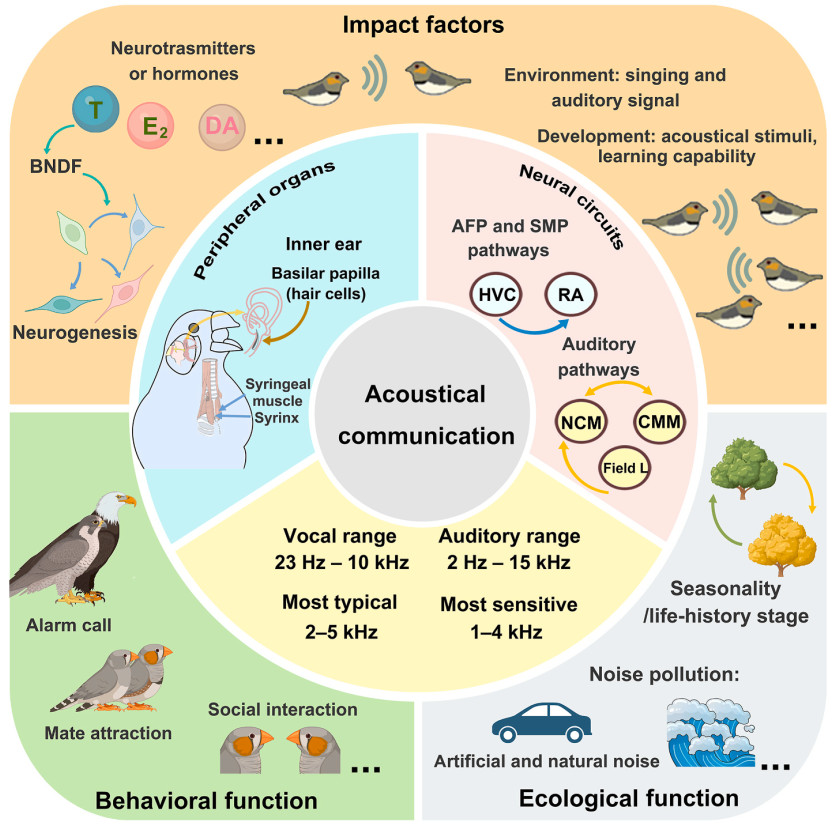





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds