Key Laboratory of Biodiversity Conservation of National Forestry and Grassland Administration, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, 100091, China
b.
Key Laboratory for Biodiversity Science and Ecological Engineering, Ministry of Education, College of Life Sciences, Beijing Normal University, Beijing, 100875, China
c.
Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China
d.
College of Life Sciences, University of Chinese Academy of Sciences, Beijing, 100049, China
e.
Qilian Mountain National Park Qinghai Province Management Bureau, Qinghai, 810400, China
Funds:
the National Natural Science Foundation of China32070530
the National Key Research and Development Program of China2019YFA0607103
the Qilian Mountain National Park Qinghai Province Management Bureau
the National Forestry and Grassland Administration of China
Key Laboratory of Forest Protection of National Forestry and Grassland Administration, National Bird Banding Center of China, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, 100091, China. E-mail address: zm7672@126.com (G. Zhang)
Peer review under the responsibility of Editorial Office of Avian Research.
Evaluating the habitat suitability of flagship species and its key influencing factors is vital for understanding potential conservation issues and developing coping strategies. We surveyed the wintering population size and distribution of the Black-necked Crane (Grus nigricollis) in the Yarlung Zangbo River Basin (YZRB) from 10 to 29 January 2022, and predicted the current potentially suitable habitat distribution and its effective factors using the MaxEnt model. A total of 9337 wintering Black-necked Cranes were recorded in the YZRB in 2022, 76.58 % of which were primarily found in Lhunzub, Samzhubze, Namling, and Lhaze. Compared to 2018, the crane population has exhibited a notable decline in Samzhubze and Taktse, likely due to farmland plowing, winter irrigation, changes in agricultural practices, road construction and hydraulic projects. The crane population within various counties exhibited a significant positive correlation with the suitable habitat area (r = 0.70, P = 0.002, n = 17). We also found that the currently suitable habitat area covered 17,204 km2, of which only 3244 km2 (18.86 %) was effectively protected at the national level, which was predominantly distributed in farmland and rangeland habitats characterized by gentle slopes, altitudes not exceeding 4500 m, and proximity to human settlements along rivers, where suitable isothermal values (51) and less seasonal precipitation (20 mm) prevail. Our study will be helpful for formulating reasonable conservation strategies to protect the core population of this threatened highland flagship species.
Habitat suitability reduction is one of the primary threats to global biodiversity. Climatic changes and human activities, such as agricultural development, urbanization, infrastructure construction, and the building of dams and reservoirs, alter land-use structures and landscape patterns, leading to the fragmentation and reduced suitability of wildlife habitats (Zhao et al., 2020; Li et al., 2022a). In the coming decades, climate change is projected to become more severe (Garcia et al., 2014), altering the global landscape and significantly impacting wildlife habitats (Parmesan and Yohe, 2003). One-sixth of extant species are estimated to be threatened with extinction due to climate-related range reductions (Urban, 2015; Xu et al., 2020). Habitat fragmentation not only restricts species movement, dispersal, and exchange but also accelerates the extinction of endangered species and exacerbates biodiversity loss (Haddad et al., 2015; Newbold et al., 2015). Focusing on flagship or umbrella species is a common strategy in regional biodiversity conservation, as it can reduce costs and enhance effectiveness (Gill et al., 2017; McGowan et al., 2020). Many studies have shown that flagship species play a key role in raising conservation funds, increasing awareness of biodiversity conservation, and maintaining ecosystem services (Wang et al., 2023). For example, in Queensland, Australia, public concern for riverine habitats significantly increased following an education and outreach campaign featuring the Platypus (Ornithorhynchus anatinus George Shaw, 1799) (Smith and Sutton, 2008). Moreover, in the western demilitarized zone of Korea, establishing a conservation plan based on the distribution of a flagship species, the White-naped Crane (Grus vipio), also protected the habitats of other threatened waterbirds (Kim et al., 2020). Strategically conserving a flagship species not only enhances its own survival prospects but also supports the protection of the entire ecosystem (Veríssimo et al., 2014). Given the increasing impact of climate change and human activities, studying the potential suitable habitat distribution range of flagship species and its influencing factors is vital for understanding potential conservation issues, developing coping strategies, and maintaining regional biodiversity.
In examinations of the habitat suitability of wildlife and its influencing factors, niche species distribution models have gained widespread popularity. Currently, the most commonly employed models include generalized linear models (GLMs), genetic algorithm for rule-set production (GARP) models, maximum entropy (MaxEnt) models, the HABITAT model, and the BIOCLIM model (Latimer et al., 2006). Each model possesses distinct advantages and limitations stemming from their underlying principles and algorithms, and their effectiveness may vary depending on the input datasets (Thuiller, 2003). The extensively utilized MaxEnt model delineates the ecological niche of target species by utilizing occurrence points and environmental data within a specific mathematical framework (Phillips and Dudík, 2008). Rooted in the principle of entropy maximization, wherein natural systems inherently maximize entropy to achieve equilibrium with their surroundings (Suárez-Seoane et al., 2008), MaxEnt conceptualizes species and their habitats as an integrated system, establishing a stable relationship by computing parameters at maximum entropy. This model predicts potential species distributions by fitting the probability distribution with the highest entropy value (Phillips et al., 2006). It requires minimal data, is user-friendly, has rapid execution, and is effective even with small sample sizes (Elith et al., 2011). Furthermore, it avoids biased assumptions concerning unknown conditions, resulting in predictions with minimal bias and high precision (Merow et al., 2013). MaxEnt has been utilized to assess suitable habitats for various avian species, including the Common Crane (Grus grus) (Stabach et al., 2009) and the Red-crowned Crane (Grus japonensis) (Wang et al., 2020).
The Black-necked Crane (Grus nigricollis), the only one of the fifteen crane species that lives its entire life on the plateau (Liu et al., 2012), is currently listed as "near threatened" by the International Union for Conservation of Nature (IUCN) Red List of Threatened Species and classified as a first-class protected bird species in China (BirdLife International, 2023). They breed on the Qinghai–Tibet Plateau and Ladakh and overwinter in low-altitude areas of the Qinghai–Tibet Plateau, Yunnan–Guizhou Plateau, Bhutan, and southern Tibet in China (Li, 2014). In particular, the Yarlung Zangbo River Basin (YZRB) of Tibet is an essential habitat for wintering Black-necked Cranes, whose numbers account for 80% of the wintering population of cranes globally. Furthermore, the Black-necked Crane is also perceived as a flagship species in alpine wetland ecosystems (Niemi et al., 1997). Therefore, research on habitat evaluation and the prediction of potentially suitable habitats for Black-necked Cranes will contribute to the development of more effective conservation strategies for protecting the wintering population of cranes and the biodiversity of the Tibetan Plateau.
The Tibetan Plateau is currently deemed a driver and an amplifier of escalating global warming, and it is a sensitive area for global climate change (Immerzeel et al., 2010; Jin et al., 2013). Moreover, with economic and social development, the intensity of human activities in the Qinghai‒Tibet Plateau, particularly in the YZRB, has gradually increased (Feng et al., 2016). In recent years, there has been a certain amount of infrastructure construction, such as highways and railways, and grassland management, such as fencing (Jia et al., 2019). Due to challenging climatic conditions and limited human resources, current systematic research targeting habitat suitability evaluation for Black-necked Cranes in the YZRB remains relatively scarce. The aims of the present study were (1) to preliminarily understand the population size and distribution of wintering Black-necked Cranes in the YZRB; (2) to determine the suitable habitat utilization types for the Black-necked Crane and explore the impact of human activities, especially agricultural development, on the population size and distribution of Black-necked Cranes; and (3) to predict the potential suitable habitat distribution range of Black-necked Cranes in the YZRB under the current climate scenario and influencing factors to formulate reasonable conservation strategies.
2.
Materials and methods
2.1
Study area
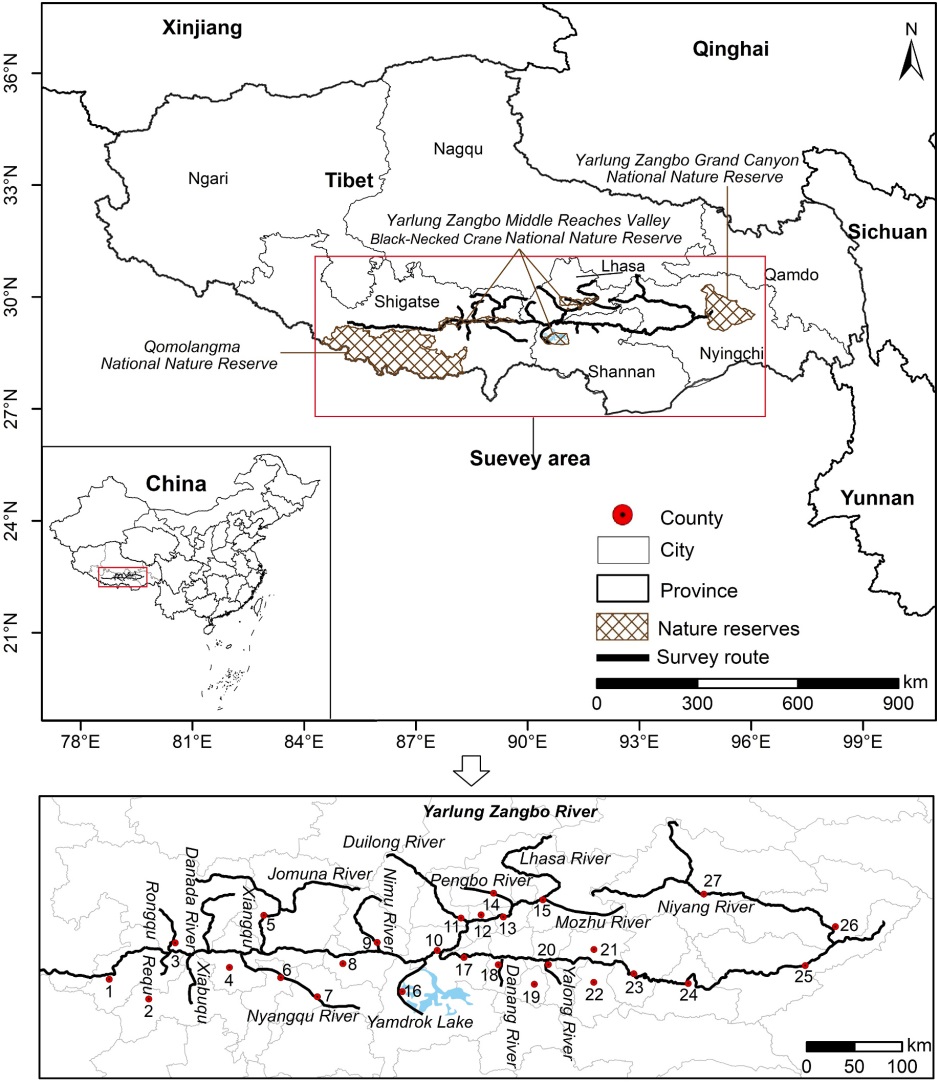
The YZRB (28°59′–30°34′ N, 82°32′–94°28′ E) is located on the southern Qinghai–Tibet Plateau in western China (Fig. 1) and has a river length of approximately 2000 km and an average elevation of 4000 m above sea level, which is affected by the combined effect of westerly and Indian monsoon winds (Yang et al., 2020). The climate is subtropical, with a mean temperature range of 5.7–10.3 ℃, an average annual precipitation range of 158.3–778.1 mm, and an annual potential evaporation range of 2293–2734 mm (Zhu et al., 2015; Yang et al., 2021). The annual average wind speed ranges from 1.6 to 3.3 m/s, and strong winds and sand flows are more common during the winter and spring (December to May), causing strong wind erosion (Yang et al., 2020).
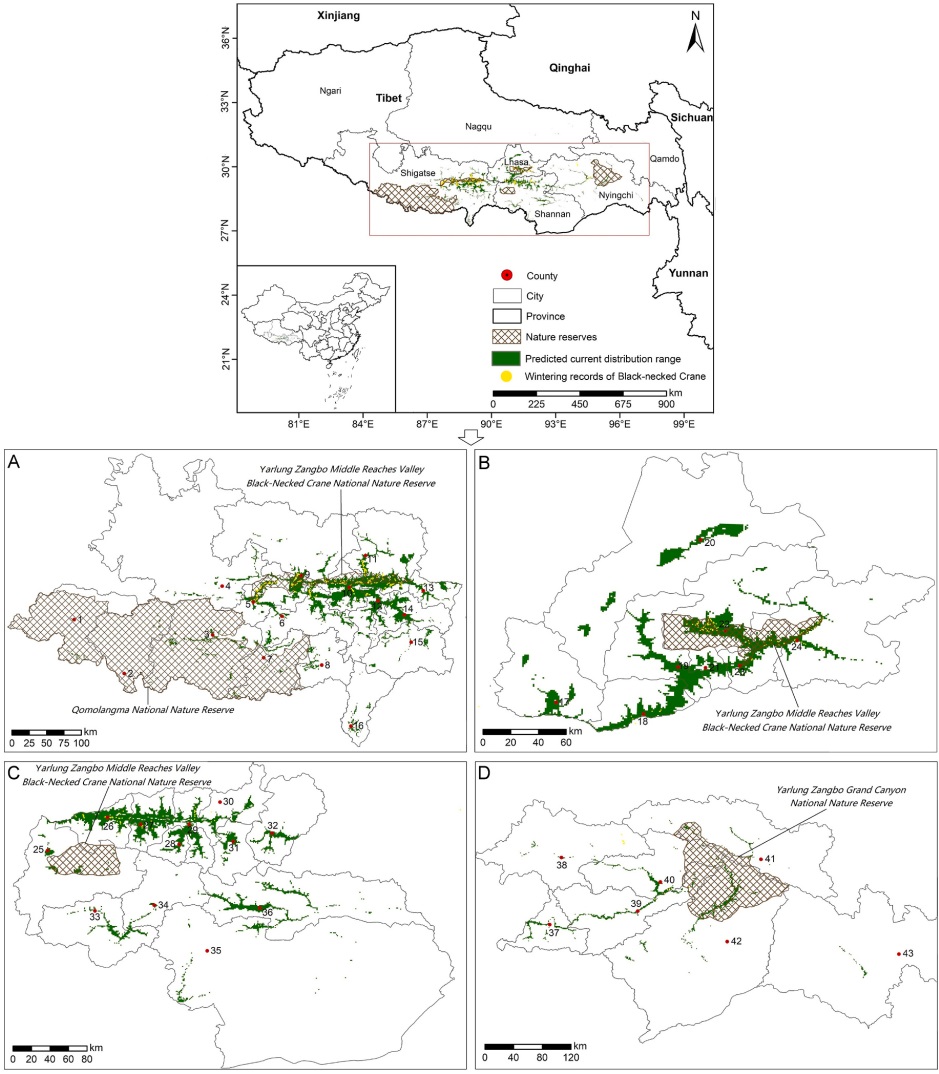
As the main agricultural area in Tibet, the YZRB is not only a local economic center but also a good habitat for the Black-necked Crane, Ruddy Shelduck (Tadorna ferruginea), Bar-headed Goose (Anser indicus), and other birds (Zhou et al., 2020). The "Yarlung Zangbo Middle Reaches Valley Black-necked Crane National Nature Reserve" has been established to strengthen the protection of wild birds, including Black-necked Cranes. Moreover, the "Qomolangma National Nature Reserve" and "Yarlung Zangbo Grand Canyon National Nature Reserve" have been established in the vicinity of the YZRB.
2.2
Methodological framework
The methodological steps in the present study were as follows: (1) surveying the population size, distribution, occurrence points and habitat types of Black-necked Cranes in the YZRB, and further comparing the population size of Black-necked Cranes in the YZRB for the years 2018 and 2022; and (2) constructing a species distribution model for wintering Black-necked Cranes in the YZRB using the MaxEnt model, based on occurrence points obtained from field surveys, climatic data from the Coupled Model Intercomparison Project Phase 6 (CMIP6), and other environmental variables.
2.3
Collection of field data
Wintering Black-necked Cranes usually arrive at the YZRB in mid-November and reach stability in early December (Jia et al., 2019). During the period of 10–29 January 2022, we conducted a survey on the population size, distribution, and habitat types of Black-necked Cranes in the YZRB. Twenty-seven counties were covered, and a total distance of approximately 1800 km was surveyed from Lhaze to Gongbo Gyamda (Fig. 1). The field survey data from 2018 is available in Jia et al. (2019).
We used 10 × binoculars to scan along the routes from 9:00 to 19:00 every day. When cranes were detected, they were observed and counted using a monocular scope (20–60 ×, SWAROVSKI, Austria), and we recorded the location, flock size, number of individuals, habitat types and presence of human disturbance, including mining development, road and railway construction, reclamation, river dredging, and presence of greenhouses for vegetable cultivation. We divided the major habitat types into winter wheat farmland, crop stubble land (including barley, rapeseed and spring wheat), plowed fields, pastures, rivers, and marshes (Jia et al., 2019). To avoid double counting, we ignored flocks flying over from behind. A flock was defined as a group of two or more cranes in close proximity and interacting with each other. To obtain the actual locations of each flock and individual crane, the positions of the observation sites were mapped with a global positioning system (GPS) device, and the distance between the cranes and surveyors was recorded using a laser range finder (ZEISS Victory 82 × 6 PRF). The direction from the cranes to the observers was measured with a compass, and then we determined the actual locations of the cranes using the polar coordinate method.
2.4
Environmental data source
We collected 19 bioclimatic variables and surface elevation data from WorldClim (http://www.worldclim.org/, accessed on 2 February 2024, v 2.1, 1970–2000). Two layers, namely, slope and aspect, were extracted from the surface elevation data using the 3D Analyst tools of ArcGIS 10.8. The population density data were downloaded from Open Spatial Demographic Data and Research (http://www.worldpop.org, accessed on 2 February 2024). The basic vector layers of rivers, lakes, roads and residential locations (villages and rural settlements) were downloaded from the Open Street Map (https://www.openstreetmap.org/, accessed on 5 February 2024). The types of roads primarily included highways, railways, and country roads. Highways were further delineated based on traffic volume, intended usage, and characteristics into expressways (annual average daily traffic (AADT) ≥ 25,000 vehicles), first-class highways (AADT = 10,000–25,000 vehicles), second-class highways (AADT = 2000–10,000 vehicles), third-class highways (AADT = 200–2000 vehicles), and fourth-class highways (AADT < 200 vehicles) (Highway Engineering Standard System (JTG 1001–2017), https://www.bengbu.gov.cn/group1/M00/00/E6/wKgQFl6dlouAAvnAAApF2o2mPDg306.pdf?). Then, four layers, namely, distance to the nearest perennial rivers, lakes, residential locations, and roads, were produced using the Euclidean Distance Tool of ArcGIS 10.8.
The land use data for Tibet, with 10 m precision, for 2022 were downloaded from the Environmental Systems Research Institute (ESRI, https://livingatlas.arcgis.com/landcoverexplorer, accessed on 2 September 2023). Subsequently, the data were imported into ArcGIS, and the "Mosaic to New Raster" tool was utilized to merge the different regional layers. Afterward, the "Extract by Mask" tool was utilized to obtain the land use layer of the study area. The ESRI GLC 2020 has nine classes (water, trees, flooded vegetation, farmland, built area, bare ground, snow/ice, clouds and rangeland) (Appendix Table S1). The monthly mean net primary productivity from June to July 2021 was obtained from the National Aeronautics and Space Administration (NASA) Earth Observation (https://neo.gsfc.nasa.gov/, accessed on 16 March 2023). The months of June and July mark the harvest season for crops, such as rapeseed, winter wheat, and highland barley, in the YZRB, which feature a unique landscape during this period, consequently allowing for more accurate measurements of net primary productivity. A complete overview and detailed information on all variables are shown in Appendix Table S2.
2.5
Habitat suitability assessment
2.5.1
Preprocessing of crane occurrence points and environmental data
We gathered 1779 wintering records of Black-necked Cranes in the YZRB areas in 2018 and 2022 via field surveys. To reduce the possibility of model overfitting, ENMTools.pl (https://github.com/danlwarren/ENMTools) software was used to trim the occurrence points so that only one observation was retained in each 30 arc-second grid cell (corresponding to the environmental variable data below) to mitigate sampling bias (Warren et al., 2010). Finally, 337 locations of the cranes remained, thus providing enough data points for constructing species distribution models (van Proosdij et al., 2016). The location data were converted into the WGS_1984_UTM_Zone_45N projected coordinate system, and decimal coordinates were calculated using the "Geometry Calculation" tool. The resulting data were stored in CSV format.
For each environmental variable, the grid size was resampled to 1 km × 1 km using a bilinear interpolation method for continuous variables and nearest neighbor assignment for categorical vegetation layers (Zhu et al., 2020). To reduce multicollinearity among the environmental variables, we used ENMTools.pl software to analyze the correlations of 35 candidate variables and removed highly correlated variables (|r| > 0.7) (Dormann et al., 2013). Eventually, 16 environmental variables, including bio3, bio7, bio14, bio15, bio19, elevation, slope, aspect, population density, d_country road, d_railway, d_settlement, d_river, d_lake, npp, and vegetation, were preserved to construct the SDMs (Appendix Table S2 and Fig. S1).
2.5.2
Optimization of model parameters
During the process of constructing distribution models, MaxEnt software automatically provides a set of system-default parameters. However, these default parameters are influenced by the test data. Models developed under default settings are susceptible to overfitting, as they are highly sensitive to the test data, which can lead to a significant discrepancy between the model's predictions and the actual distribution. Consequently, the optimization of model parameters is imperative for enhancing the accuracy of predictions and the reliability of the outcomes (Fernández and Morales, 2019).
The regularization multiplier and combination of features may influence the predictive performance and accuracy of the model (Ji et al., 2021), which are essential for building a species distribution model using MaxEnt 3.4.4 software. Therefore, we optimized these two parameters by using the Kuenm (https://github.com/marlonecobos/kuenm) package in R 3.6.3 (https://www.r-project.org/) (Cobos et al., 2019). In the modeling, 75% of the data were used as the training set. Overall, 1160 candidate models, with parameters reflecting all combinations of 40 regularization multiplier settings (from 0.1 to 4, with an interval of 0.1), and 29 feature class combinations [the MaxEnt software provides five feature classes: L (linear), Q (quadratic), P (product), H (hinge), T (threshold)], were evaluated (Li et al., 2023). Model selection was based on statistical significance (partial ROC), predictive ability (low omission rates), and complexity [Akaike's Information Corrected Criterion (AICc) values], in that order of priority (Li et al., 2023). First, candidate models were screened to keep those that were statistically significant; second, the set of models was reduced with the omission rate criterion (i.e., < 5% when possible); finally, the models with the lowest delta AICc values (<2) were selected among the significant and low-omission candidate models (Li et al., 2023). Consequently, the model whose feature combination and regularization multiplier were L + 0.1 was used to construct a MaxEnt model for the current Black-necked Crane range (Appendix Fig. S2).
2.5.3
Construction of the species distribution model
We built the current species distribution model with MaxEnt 3.4.4. All the remaining Black-necked Crane distribution points were imported into MaxEnt software, 75 % of the geographic distribution data were used for training, and 25% were used to validate the model (Li et al., 2022a, Li et al., 2023). The output format was "Logistic", the output file type was "ASCII", the maximum iteration mode was set to "Bootstrap", and the number of repetitions was 10. To determine the importance of each environmental variable, the jackknife test was performed (Phillips et al., 2006). The AUC was selected to evaluate the model; the output values of the model were continuous (Xu et al., 2015). The AUC was indicated with the mean values of the training and test AUC values of 10 model replicates. The probability that the output value of the model in each grid cell is between 0 and 1 can be interpreted as relative suitability. We used the maximum test sensitivity plus specificity (MTSPS) threshold to reclassify the model outputs: grid cells with suitability values higher than the MTSPS threshold were regarded as potentially suitable areas, and other grid cells were regarded as unsuitable areas. In this study, the MTSPS was 0.1436.
Finally, the ASC file (average value of 10 runs) obtained by running the MaxEnt model was imported into ArcGIS 10.8. The "Reclassify" tool, with a threshold of 0.1436, was used to obtain suitable habitats (MTSPS ≥ 0.1436) and unsuitable habitats (MTSPS < 0.1436) for Black-necked Cranes. Then, the vector boundary data of protected areas and the boundary data of each county in Tibet were used, and the "Extract by Mask" tool was used to obtain the area of protected suitable habitats and the area of suitable habitats in each county and calculate their proportions.
2.6
Data analysis
We counted the number of Black-necked Cranes in each county in 2022 and compared it with that in 2018, analyzing the changes in the number of cranes in each county. Three clustering patterns of Black-necked Cranes were commonly observed: family cranes, flocking cranes, and occasionally solitary cranes. A family group typically comprised 1–2 adults and ≤2 juveniles. Furthermore, for the purposes of data collection and analysis, flocking cranes with an individual count exceeding four were categorized into four groups: 5–10 individuals, 11–50 individuals, 51–100 individuals, and > 100 individuals. The normality of the data regarding the average group size of Black-necked Cranes across different habitat types was assessed using the Kolmogorov‒Smirnov (K–S) test, which confirmed a normal distribution. Consequently, one-way analysis of variance (one-way ANOVA) was employed to evaluate whether significant differences existed in the average group sizes of Black-necked Cranes among the different habitat types, followed by a least significant difference (LSD) test for multiple pairwise comparisons. The Pearson correlation coefficient was used to analyze the relationships between the number of Black-necked Cranes in various administrative regions and the area of suitable habitats within those regions.
3.
Results
3.1
Population size and habitat use
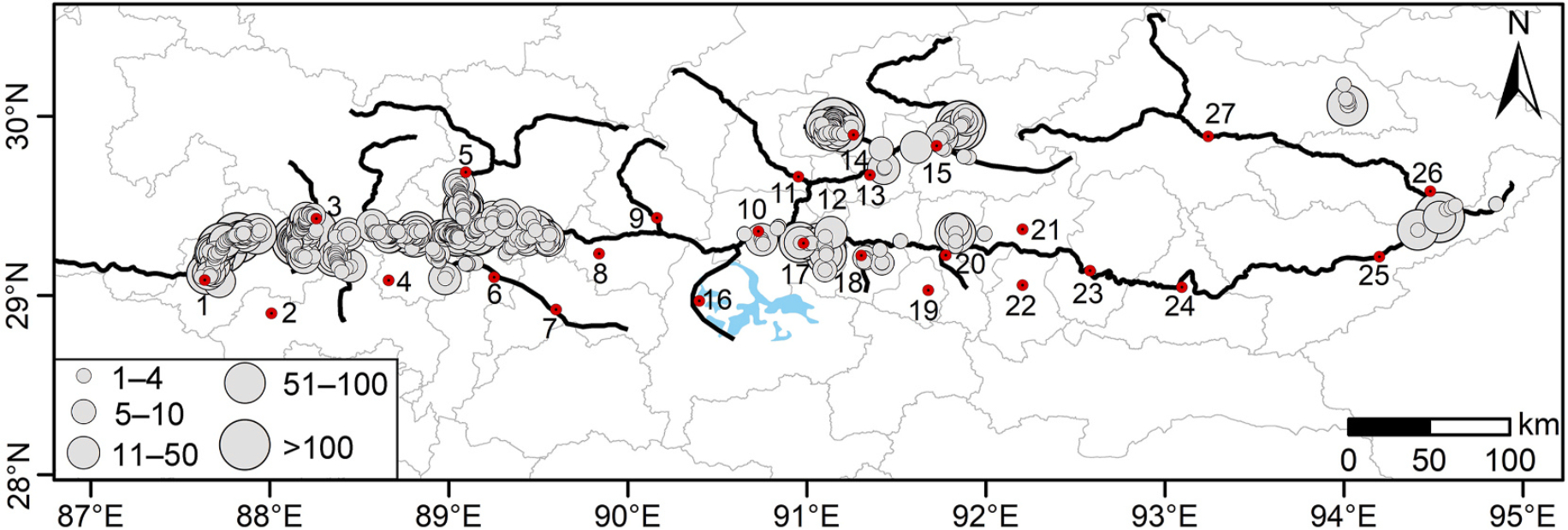
A total of 9337 Black-necked Cranes were recorded during the survey in the middle wintering period, 2022, and the sites with the most cranes (>1000 individuals) included Lhunzub, Samzhubze, Namling, and Lhaze (Table 1, Fig. 2). Overall, there was an increase of 654 cranes in the YZRB in 2022 compared to that recorded in 2018 (the number of Black-necked Cranes in Nyingchi was not included in the comparison) (Table 1). Specifically, large increases were observed at Lhunzub, Gonggar, Namling, and Lhaze (approximately 300–500 individuals), followed by Medro Gongkar and Tongmon (approximately 150 individuals), but large decreases were observed at Samzhubze and Taktse (approximately 500–1000 individuals) (Table 1). Moreover, a total of 1288 flocks were recorded, with an average flock size of 7.25 ± 0.50 cranes (n = 1288, range 1–208), of which the number of single families accounted for 73.60%, followed by flocks with 5–10 and 11–50 individuals, accounting for 7.92% and 7.84%, respectively (Fig. 2).
Table
1.
The number and distribution of Black-necked Cranes in the Yarlung Zangbo River Basin, Tibet, 2018 and 2022.
City
County
Middle wintering period in 2022
Middle wintering period in 2018a
Shigatse
Lhaze
2090
1711
Sakya
212
236
Thongmon
468
308
Samzhubze
1780
2735
Namling
1943
1366
Panam
22
66
Gyantse
0
15
Rinbung
0
1
Subtotal
6515
6438
Lhasa
Quxu
14
17
Taktse
33
574
Lhunzub
1337
830
Medro Gongkar
353
210
Subtotal
1739
1631
Shannan
Gonggar
528
118
Danang
39
2
Nedong
122
63
Sangri
4
39
Subtotal
693
222
Nyingchi
Gongbo Gyamda
86
–
Bayi
10
–
Mainling
296
–
Subtotal
392
–
Total
9337
8291
a Indicates that the data were obtained from Jia et al. (2019); numbers in bold indicates the change of individual numbers between 2018 and 2022 greater than 300; "–" means data not available.
Figure
2.
Distribution of wintering Black-necked Crane survey sites in the Yarlung Zangpo River Basin, Tibet, 2022. The gray points indicate the population size of Black-necked Cranes recorded via a single survey in that area. The place names represented by numbers 1–27 are the same as those in Fig. 1.
Crop stubble land and plowed land were the main foraging habitats for Black-necked Cranes during the middle wintering period, and the numbers of individuals (flocks) in these two habitats accounted for 58.18% (52.62%) and 21.61% (27.91%) of the total observed cranes, respectively (Table 2). Crane flocks of different sizes exhibited variations in habitat utilization (F5,1271 = 4.67, P < 0.001); the average flock size of cranes recorded in the river was larger than that in the pasture (P < 0.001), winter wheat land (P = 0.004), plowed land (P < 0.001), crop stubble land (P < 0.001), and marsh (P < 0.001); the average flock size of cranes recorded in the crop stubble land was larger than that in the plowed land (P = 0.004); and there were no significant differences in the average flock sizes of cranes across the other habitats (P > 0.05) (Table 2).
Table
2.
The number of Black-necked Cranes in different habitats in the Yarlung Zangbo River Basin, Tibet, 2022.
3.2
The main environmental variables affecting the habitat suitability
The average training AUC and test AUC for the model presented in Section 2.5.3 were 0.981 and 0.980, respectively, which indicated that the MaxEnt model for the current distribution range of the Black-necked Cranes performed well (Appendix Fig. S3). The contribution of each environmental variable to the model showed that vegetation contributed 32.2% to the model, followed by d_railway (25.8%), elevation (12.3%), bio3 (9.86%), bio15 (6.73%), d_settlement (4.3%), d_river (4.1%), and slope (2.7%). These eight predictors contributed 97.91% to the model (Appendix Table S3).
In terms of habitat, Black-necked Cranes prefer farmland and rangeland habitats with gentle slopes along rivers, situated at altitudes not exceeding 4500 m, often near human settlements (Fig. 3A–C, F–H); in terms of hydrothermal conditions, they prefer relatively isothermally suitable areas (bio3) (51) (Fig. 3D) and smaller areas of precipitation that vary seasonality (bio15) (20 mm) (Fig. 3E).
Figure
3.
Response curves of habitat suitability for the Black-necked Crane (vertical axis) to vegetation (A), d_railway (B), elevation (C), bio3 (D), bio15(E), d_settlement (F), d_river (G), and slope (H), with standard deviation (SD, blue buffers).
3.3
Current suitable habitat ranges and protection status
According to model predictions, the suitable area for Black-necked Cranes in the YZRB under current climate conditions was 17,204 km2, of which only 3244 km2 was effectively protected, accounting for 18.86%, and a considerable amount of potentially appropriate habitats remained unprotected. These suitable habitats were primarily distributed along the middle reaches of the YZRB and its tributaries. Samzhubze has the largest area of suitable habitat for Black-necked Cranes (accounting for 10.32%), followed by 25 other counties, each with suitable habitat areas exceeding 250 km2 (Fig. 4, Appendix Table S4). The suitable habitat range of Black-necked Cranes in the YZRB spans longitudinally to the west of Ngamring and Jilong of Shigatse and to the east of Bomi and Zayv of Nyingchi, latitudinally to the north of Damxung of Lhasa, and the range extends to the southern periphery, encompassing Yadong of Shigatse and Cuona of Shannan (Fig. 4).
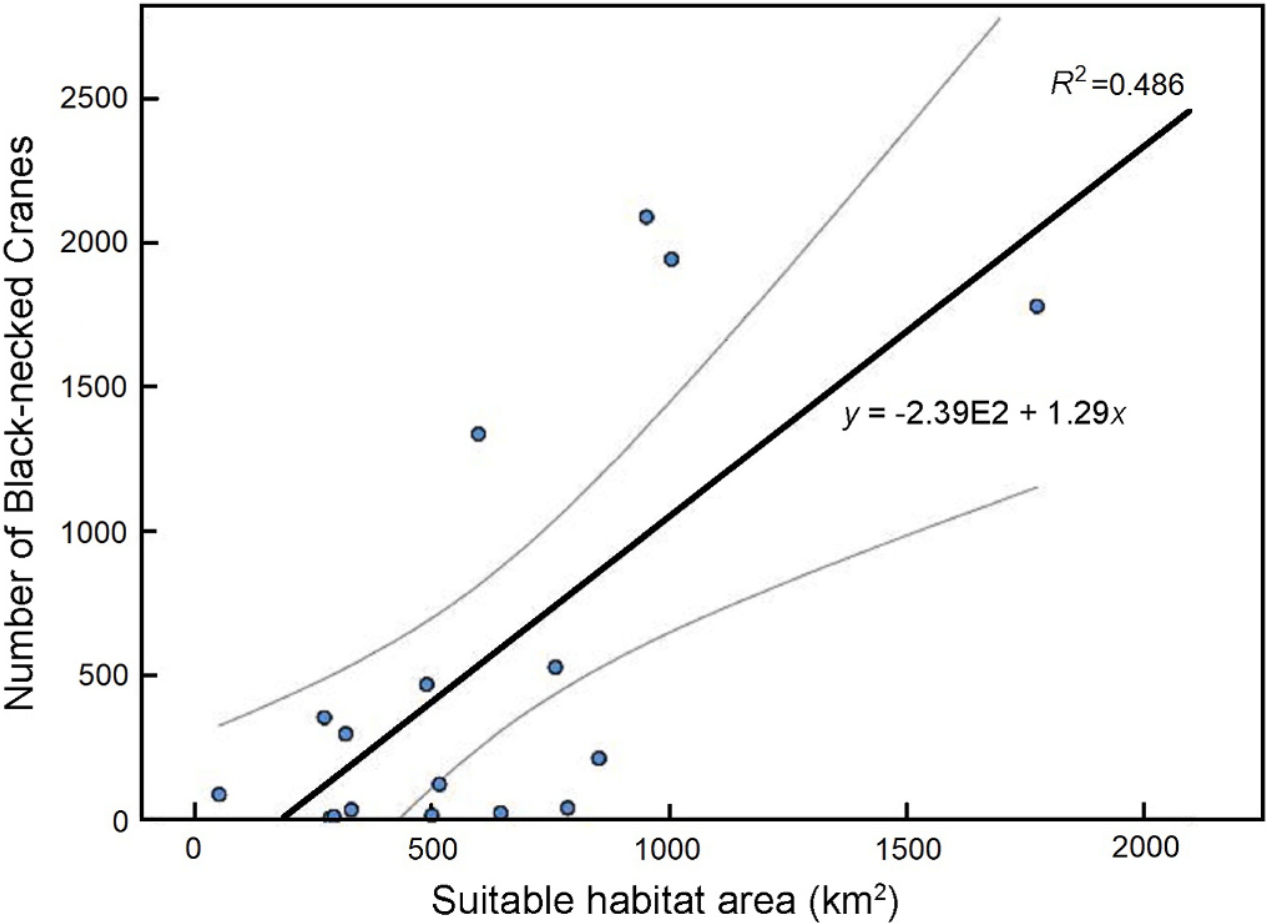
Moreover, we discovered that the number of Black-necked Cranes across various counties significantly increased as the suitable habitat area increased (r = 0.70, P = 0.002, n = 17) (Fig. 5).
Figure
5.
The correlation between the number of Black-necked Cranes and the area of their suitable habitats.
In this study, we confirmed the population size and spatial distribution of the wintering Black-necked Cranes in the YZRB, identified the main climatic and environmental variables impacting habitat suitability for Black-necked Cranes, and predicted the current potential distribution range and conservation gaps in suitable habitats within the YZRB. Finally, we proposed pertinent conservation strategies for the future protection and management of wintering Black-necked Crane populations in the YZRB.
4.1
Changes in the population size and distribution
In this survey, a cumulative number of 9337 Black-necked Cranes was documented within the stretch extending from Lhaze to Gongbo Gyamda. Notably, the cumulative number of cranes in Lhaze, Namling, Samzhubze, Lhunzub, and Gonggar accounted for 76.58% of the observed population. These five counties were the critical wintering habitats for the Black-necked Crane in the YZRB, which was consistent with the results previously reported by Yang et al. (2016). Predictions from the MaxEnt model also indicated that these five counties possessed larger suitable habitat areas for the cranes (Appendix Table S4). Furthermore, in comparison to that in 2018, the number of Black-necked Cranes significantly increased in the YZRB in 2022 (Jia et al., 2019). Notably, there was a significant increase in Lhunzub, Gonggar, Namling, and Lhaze; however, there was a significant decrease in Samzhubze and Taktse (Jia et al., 2019). Landscape change and fragmentation of suitable habitats (including rangeland, farmland, and water) due to anthropogenic disturbances are significant factors affecting the population size and distribution of Black-necked Cranes (Yang et al., 2016, Yang et al., 2023). For the four counties where the number of Black-necked Cranes has increased, we speculated that this finding may be related to the increase in the area of suitable habitats for Black-necked Cranes, such as water, rangeland, wetland, and farmland. For instance, in recent years, Lhunzub has experienced a rise in wetland water levels and an expansion of marshy grassland areas, attributed to local governmental initiatives in wetland conservation and rehabilitation, including the establishment of protective forest buffers, controlled grazing practices, and measures for wetland hydrological management and augmentation (Wang et al., 2021).
However, regarding the reasons for the decline in the number of Black-necked Cranes in Samzhubze and Taktse, we speculated that the decrease in the number of Black-necked Cranes in these areas may be related to land cultivation and winter irrigation activities. Field surveys indicated that post-harvest remnants of highland barley, indigenous grass species, and emergent winter wheat crops constituted the primary sources of Black-necked Cranes during the colder months in the YZRB. More than 50% of the cranes exhibited a preference for foraging on uncultivated crop stubble land (Table 2). With the onset of more severe winter conditions, the farmland was plowed and irrigated by farmers, which was essential for soil moisture retention and the facilitation of seed germination and subsequent barley cultivation in the ensuing growing season. However, agricultural tillage may result in the deposition of seeds deep within the soil strata and ice from winter irrigation, thereby decreasing the availability of foraging resources for Black-necked Cranes in cultivated fields. In contrast, stubble fields offer more available food that is easier to obtain (Bishop and Li, 2002). Consequently, many Black-necked Cranes gather on unplowed farmland, forming a sizable wintering population. Field surveys revealed that in January, there was a substantial escalation in the amount of arable land being plowed in Samzhubze. This surge has exacerbated the foraging challenges for Black-necked Cranes, compelling them to migrate to the adjacent Namling region in search of sustenance. These findings underscore the critical importance of farmlands to the survival of the overwintering Black-necked Cranes in the YZRB (Wu et al., 2020), corroborating the findings of Sun et al. (2018) on the foraging habitat types of the overwintering Black-necked Cranes in Caohai, Guizhou.
Moreover, recent shifts in Tibet's agricultural development model, road construction, and implementation of hydraulic projects have posed severe threats to the available habitat for overwintering Black-necked Cranes (Jia et al., 2019). For example, many greenhouses have been erected in the farmlands of the eastern YZRB, particularly in Quxu, to cultivate melons and vegetables. The construction of expressways from Lhasa to Nyingchi and Shigatse disrupted the foraging grounds of Black-necked Cranes in Quxu (Wu, 2011). The rapid development of mining in Sakya has led to an increase in human activity and industrial wastewater, which has degraded the water quality of the YZRB; dredging projects in the Lhasa River and the YZRB have directly destroyed Black-necked Crane habitats, especially its feeding grounds (Jia et al., 2019). In recent years, the proportion of forestland, grassland, and arable land in Tolun Dechen has changed due to the increase in industrial and infrastructure land associated with urbanization, as well as the implementation of policies, such as converting farmland back to forest or grassland (Liu et al., 2019).
4.2
Main environmental variables affecting the habitat suitability
In this study, vegetation, d_railway, elevation, bio3, bio15, d_settlement, d_river, and slope were identified as the primary environmental variables influencing habitat suitability for Black-necked Cranes. The MaxEnt model's predictive outcomes indicated that agricultural land exhibited the greatest habitat suitability for Black-necked Cranes, which was consistent with the results of our field investigation. Black-necked Cranes wintering in the Dashanbao National Nature Reserve in Yunnan Province (Wu et al., 2021) and Caohai in Guizhou Province (Wu et al., 2020) also exhibited a preference for agricultural landscapes. In addition, other crane species, such as Sarus Cranes (Grus antigone) (Mukherjee et al., 2002) and Brolgas (Grus rubicunda) (Sheldon, 2005), also show a preference for agricultural landscapes. Furthermore, the curve showing the influence of the altitudinal factor indicated that Black-necked Cranes avoid areas with altitudes above 4500 m and slopes steeper than 15°, closely matching the results (2882–4256 m) from field surveys in the YZRB. Studies have shown that the altitudinal gradient on the Tibetan Plateau, accompanied by variations in temperature and precipitation, acts as a natural barrier to species dispersal (Wang et al., 2004). With increasing altitude, decreasing temperature, and reduced living space and food resources, the environment becomes less favorable for the survival of Black-necked Cranes.
The middle reaches of the YZRB not only serve as concentrated areas of farmland but also act as hubs for political, economic, and cultural activities, with 96% of urban and rural land use focused along the river system, which thus experiences significant human disturbance (Li et al., 2022b). In this study, the MaxEnt model revealed that the Black-necked Crane exhibited a preference for habitats near anthropogenic settlements, indicating that the crane has not been significantly impacted by the juxtaposition of foraging environments and human disturbances, thereby demonstrating a notable level of tolerance to anthropogenic activities. This was presumed to be due to the heightened awareness of Black-necked Crane conservation among local residents in recent years, which has allowed the cranes to appear in areas of intense human activity. Similar findings were reported by Ma et al. (2021) in his research on the habitat suitability of the Crested Ibis (Nipponia nippon) under climate change conditions, wherein the species was observed to recolonize regions with intense anthropogenic activities, a phenomenon attributed to heightened conservation consciousness among residents of the Qinba Mountains. Other birds, such as the Black-headed Ibis (Threskiornis melanocephalus), also maintain habitats in highly disturbed urban areas (Koli et al., 2019). Moreover, studies by Li and Li (2005) revealed that Black-necked Cranes could share wetland resources with local residents and coexist harmoniously with humans under conditions of moderate population density and low disturbance levels. Liu et al. (2006) reported that Black-necked Cranes tend to forage around farmlands adjacent to country roads and residential areas, which typically yield the highest crop production and contain abundant crop residues. Our current research findings corroborate these historical studies. Through our field investigations, we also observed that Black-necked Cranes frequently forage in crop stubble, plowed fields, and winter wheat fields near residential areas (Appendix Fig. S4). Therefore, strengthening local residents' awareness of Black-necked Cranes conservation has been an effective measure for promoting the protection of this species and other rare birds in the YZRB, which has contributed to the harmonious coexistence of humans and avifauna.
4.3
Conservation gaps and recommendations
Only 3244 km2 of land, accounting for 18.86% of the suitable habitat for wintering Black-necked Cranes, is currently protected by the existing conservation network in the YZRB. Compared with those in 1994 (2626 km2), 2005 (2897 km2), and 2015 (2903 km2), the areas of suitable habitat of Black-necked Cranes that were protected increased from 1994 to 2022 (Yan et al., 2018). However, there are still many suitable habitats for cranes outside nature reserves, which may be associated with decreases in habitat suitability because of climate change and anthropogenic activities (Hoffmann et al., 2019; Elsen et al., 2020). Expanding and enhancing networks of protected areas are at the forefront of efforts to conserve and restore global biodiversity, but climate change and habitat loss can interact synergistically to undermine the potential benefits of networks of protected areas (Asamoah et al., 2021).
To better protect Black-necked Cranes and their habitats, we suggest the following for the conservation of this local population: (1) many suitable habitats for Black-necked Cranes lie outside the current protected area, while the current national nature reserves include unsuitable urban areas and mountainous regions. Thus, it is recommended to revise the boundaries of the YZRB reserve, specifically expanding south of Shigatse and Lhasa and north of Shannan, to better protect suitable crane habitats. (2) More than 50% of Black-necked Cranes in the YZRB rely on crop stubble land for foraging. Local governments should coordinate with relevant departments to reduce pesticide application, adjusting plowing and irrigation schedules to minimize human disturbances. Especially regarding plowing farmland and winter irrigation, reasonable rotation programs should be implemented to avoid simultaneous plowing and winter irrigation in areas where Black-necked Cranes are concentrated. (3) Rangeland, which serves as a secondary foraging habitat for Black-necked Cranes, necessitates the implementation of sustainable grazing practices. These practices include seasonal grazing prohibitions and the regulation of stocking densities to mitigate grassland degradation while simultaneously supporting pastoral livelihoods and conserving critical habitats. (4) To ensure suitable climate conditions for Black-necked Cranes, a long-term monitoring system is necessary to assess the impact of temperature and rainfall changes on wetland water levels and food supplies. In climate-vulnerable areas such as like high-altitude habitats, proactively planning ecological corridors will enhance habitat connectivity and help cranes adapt to new migration patterns.
5.
Conclusions
In this study, by integrating field survey data, we conducted a comprehensive assessment of the population size and distribution, habitat use, and key factors influencing potentially suitable habitats for Black-necked Cranes in the YZRB. Our findings provided crucial scientific insights that will guide the formulation of more effective conservation strategies. This information is vital not only for the protection of Black-necked Cranes, a critical species in the plateau ecosystem, but also to set a standard for conservation initiatives targeting other endangered bird species. Future studies should extend these predictions to include potential suitable habitat ranges under various climate change scenarios, thereby providing critical scientific support for crane conservation planning in the context of anticipated environmental changes.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We acknowledge the contributions of specific colleagues, institutions, or agencies that aided the efforts of the authors. We also thank the two anonymous reviewers for their helpful comments and suggestions that greatly improve this paper.
Asamoah, E.F., Beaumont, L.J., Maina, J.M., 2021. Climate and land-use changes reduce the benefits of terrestrial protected areas. Nat. Clim. Change 11, 1105–1110. .
BirdLife International, 2023. Grus nigricollis. The IUCN Red List of Threatened Species 2020: e. T22692162A180030167. . (Accessed 2 September 2023).
Bishop, M.A., Li, F.S., 2002. Effects of farming practices in Tibet on wintering black-necked crane (Grus nigricollis) diet and food availability. Biodivers. Sci. 10, 393–398. . (in Chinese).
Cobos, M.E., Peterson, A.T., Barve, N., Osorio-Olvera, L., 2019. kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ 7, e6281. .
Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al., 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. .
Elith, J., Phillips, S.J., Hastie, T.J., Dudík, M., Chee, Y.E., Yates, C.J., 2011. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17, 43–57. .
Elsen, P.R., Monahan, W.B., Dougherty, E.R., Merenlender, A.M., 2020. Keeping pace with climate change in global terrestrial protected areas. Sci. Adv. 6, eaay0814. .
Feng, F.F., Yang, Z.S., Owens, J.R., Hou, R., Zhang, Z.H., Qi, D.W., 2016. An assessment of endangered species habitat at large scale: chiru distribution across the Tibetan region of Chang Tang. Folia Zool. 65, 65–71. .
Fernández, I.C., Morales, N.S., 2019. One-class land-cover classification using MaxEnt: the effect of modelling parameterization on classification accuracy. PeerJ 7, e7016. .
Garcia, R.A., Cabeza, M., Rahbek, C., Araujo, M.B., 2014. Multiple dimensions of climate change and their implications for biodiversity. Science 344, 486. .
Gill, D.A., Mascia, M.B., Ahmadia, G.N., Glew, L., Lester, S.E., Barnes, M., et al., 2017. Capacity shortfalls hinder the performance of marine protected areas globally. Nature 543, 665–669. .
Haddad, N.M., Brudvig, L.A., Clobert, J., Davies, K.F., Gonzalez, A., Holt, R.D., et al., 2015. Habitat fragmentation and its lasting impact on Earth's ecosystems. Sci. Adv. 1, e1500052. .
Hoffmann, S., Irl, S.D.H., Beierkuhnlein, C., 2019. Predicted climate shifts within terrestrial protected areas worldwide. Nat. Commun. 10, 4787. .
Immerzeel, W.W., van Beek, L.P.H., Bierkens, M.F.P., 2010. Climate change will affect the Asian water towers. Science 328, 1382–1385. .
Ji, W., Gao, G., Wei, J., 2021. Potential global distribution of Daktulosphaira vitifoliae under climate change based on MaxEnt. Insects 12, 347. .
Jia, R., Ma, T., Zhang, F.J., Zhang, G.G., Liu, D.P., Lu, J., 2019. Population dynamics and habitat use of the black-necked crane (Grus nigricollis) in the Yarlung Tsangpo River basin, Tibet, China. Avian Res. 10, 32. .
Jin, Z.N., Zhuang, Q.L., He, J.S., Luo, T.X., Shi, Y., 2013. Phenology shift from 1989 to 2008 on the Tibetan Plateau: an analysis with a process-based soil physical model and remote sensing data. Clim. Change 119, 435–449. .
Kim, J.H., Park, S., Kim, S.H., Lee, E.J., 2020. Identifying high-priority conservation areas for endangered waterbirds using a flagship species in the Korean DMZ. Ecol. Eng. 159, 106010. .
Koli, V.K., Chaudhary, S., Sundar, K.S.G., 2019. Roosting ecology of black-headed ibis (Threskiornis melanocephalus) in urban and rural areas of southern Rajasthan, India. Waterbirds 42, 51–60. .
Latimer, A.M., Wu, S.S., Gelfand, A.E., Silander Jr, J.A., 2006. Building statistical models to analyze species distributions. Ecol. Appl. 16, 33–50. .
Li, D.X., Li, Z.X., Liu, Z.W., Yang, Y.J., Khoso, A.G., Wang, L., Liu, D.G., 2023. Climate change simulations revealed potentially drastic shifts in insect community structure and crop yields in China's farmland. J. Pest. Sci. 96, 55–69. .
Li, M.M., Zhou, H.M., Bai, J., Zhang, T.X., Liu, Y.X., Ran, J.H., 2022a. Distribution of breeding population and predicting future habitat under climate change of blacknecked crane (Grus nigricollis Przevalski, 1876) in Shaluli Mountains. Animals 12, 2594. .
Li, P.J., Zuo, D.P., Xu, Z.X., Gao, X.X., 2022b. Land use/cover and landscape patterns based on terrain in the Yarlung Tsangpo River basin, China. Mt. Res. 40 (1), 136–150. (in Chinese).
Li, Z.M., Li, F.S., 2005. Research on Black-Necked Cranes (Grus nigricollis). Shanghai Scientific and Technological Education. Publishing House, Shanghai, China.
Liu, N., Kuang, F.L., Xia, F., Zhong, X.Y., 2006. Aprimary study on feeding habitat use of black-necked crane during winter. J. West China For. Sci. 35, 25–32. . (in Chinese).
Liu, Q., Li, F.S., Buzzard, P., Qian, F., Zhang, F., Zhao, J.L., Yang, J.X., Yang, X.J., 2012. Migration routes and new breeding areas of black-necked cranes (Grus nigricollis). Wilson J. Ornithol. 124, 704–712. .
Liu, S.S., Yixi, D., Yang, L., Zhou, S.L., 2019. Changes in land use types may affect the distribution of wintering black-necked cranes (Grus nigricollis): a case study of Duilongdeqing District, Lhasa. Xizang Sci. Technol. 6, 11–14. . (in Chinese).
Ma, L.M., Li, X.H., Zhai, T.Q., Zhang, Y.Z., Song, K., Holyoak, M., et al., 2021. Changes in the habitat preference of crested ibis (Nipponia nippon) during a period of rapid population increase. Animals 11, 2626. .
McGowan, J., Beaumont, L.J., Smith, R.J., Chauvenet, A.L.M., Harcourt, R., Atkinson, S.C., et al., 2020. Conservation prioritization can resolve the flagship species conundrum. Nat. Commun. 11, 994. .
Merow, C., Smith, M.J., Silander Jr, J.A., 2013. A practical guide to MaxEnt for modeling species' distributions: what it does, and why inputs and settings matter. Ecography 36, 1058–1069. .
Mukherjee, A., Borad, C.K., Parasharya, B.M., 2002. Breeding performance of the Indian sarus crane (Grus antigone) in the agricultural landscape of western India. Biol. Conserv. 105, 263–269. .
Newbold, T., Hudson, L.N., Hill, S.L.L., Contu, S., Lysenko, I., Senior, R.A., et al., 2015. Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50. .
Niemi, G.J., Hanowski, J.M., Lima, A.R., Nicholls, T., Weiland, N., 1997. A critical analysis on the use of indicator species in management. J. Wildl. Manag. 61, 1240–1252. .
Parmesan, C., Yohe, G., 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42. .
Phillips, S.J., Anderson, R.P., Schapire, R.E., 2006. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. .
Phillips, S.J., Dudík, M., 2008. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31, 161–175. .
Sheldon, R.A., 2005. Breeding and flocking: comparison of seasonal wetland habitat use by the brolga (Grus rubicunda) in South-western Victoria. Aust. Field Ornithol. 22, 5–11. .
Smith, A.M., Sutton, S.G., 2008. The role of a flagship species in the formation of conservation intentions. Hum. Dimens. Wildl. 13, 127–140. .
Stabach, J.A., Laporte, N., Olupot, W., 2009. Modeling habitat suitability for grey crowned-cranes (Balearica regulorum gibbericeps) throughout Uganda. Int. J. Biodivers. Conserv. 1, 177–186. .
Suárez-Seoane, S., de la Morena, E.L.G., Prieto, M.B.M., Osborne, P.E., de Juana, E., 2008. Maximum entropy niche-based modelling of seasonal changes in little bustard (Tetrax tetrax) distribution. Ecol. Model. 219, 17–29. .
Sun, X.J., Zhang, M.M., Hannah, L., Hu, C.S., Su, H.J., 2018. Field observations on the behavior of wintering black-necked cranes (Grus nigricollis) at roosting sites in Caohai, Guizhou. Chin. J. Zool. 53 (2), 180–190. . (in Chinese).
Thuiller, W., 2003. Biomod-optimizing predictions of species distributions and projecting potential future shifts under global change. Glob. Change Biol. 9, 1353–1362. .
van Proosdij, A.S.J., Sosef, M.S.M., Marc, S.M., Wieringa, J.J., Raes, N., 2016. Minimum required number of specimen records to develop accurate species distribution models. Ecography 39, 542–552. .
Veríssimo, D., Pongiluppi, T., Santos, M.C., Develey, P.F., Fraser, I., Smith, R.J., et al., 2014. Using a systematic approach to select flagship species for bird conservation. Conserv. Biol. 28, 269–277. .
Wang, G., Wang, C., Guo, Z.R., Dai, L.J., Wu, Y.Q., Liu, H.Y., et al., 2020. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: suggestions for conservation of endangered red-crowned crane (Grus japonensis). Ecol. Indic. 116, 106472. .
Wang, L., Ma, L.R., Xu, Z.G., Xie, Y.F., 2021. Protection and restoration of ganqu wetland in linzhou county. J. Green Sci. Technol. 23 (16), 162–165. . (in Chinese).
Wang, Y.F., Shi, J., Wu, Y., Zhang, W.W., Yang, X., Lv, H.X., et al., 2023. Selection of flagship species and their use as umbrellas in bird conservation: a case study in Lishui, Zhejiang Province, China. Animals 13, 1825. .
Wang, Z.H., Chen, A.P., Piao, S.L., Fang, J.Y., 2004. Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China. Biodivers. Sci. 82–88. (in Chinese).
Warren, D.L., Glor, R.E., Turelli, M., 2010. ENMTools: a toolbox for comparative studies of environmental niche models. Ecography 33, 607–611. .
Wu, D.W., Hu, C.S., Zhang, M.M., Li, Z.M., Su, H.J., 2020. Foraging habitat selection of overwintering black-necked cranes (Grus nigricollis) in the farming area surrounding the Caohai Wetland, Guizhou Province, China. Avian Res. 11, 5. .
Wu, F.Z., Wang, J.L., Shen, J., Hu, W.Y., Zheng, M.H., Dao, M.B., 2021. A study on the suitability evaluation of the black-necked crane foraging area in Dashanbao Nature Reserve, Yunnan Province. Geomatics Spatial Inf. Technol. 44 (8), 96–100. . (in Chinese).
Wu, Y., 2011. Based on the environmental impact assessment of ecological behavior analysis. Railw. Energy Sav. Environ. Prot. Occup. Saf. Health. 1 (4), 200–205. . (in Chinese).
Xu, Y., Wang, B., Zhong, X., Yang, B., Zhang, J.D., Zhao, C., et al., 2020. Predicting range shifts of the Chinese monal (Lophophorus lhuysii) under climate change: implications for long-term conservation. Glob. Ecol. Conserv. 22, e01018. .
Xu, Z.L., Peng, H.H., Peng, S.Z., 2015. The development and evaluation of species distribution models. Acta Ecol. Sin. 35, 557–567. .
Yang, J.H., Xia, D.S., Gao, F.Y., Wang, S.Y., Chen, Z.X., Jia, J., et al., 2020. Aeolian deposits in the Yarlung Zangbo River Basin, southern Tibetan plateau: a brief review. Adv. Earth Sci. 35 (8), 863–877. . (in Chinese).
Yang, J.H., Xia, D.S., Gao, F.Y., Wang, S.Y., Li, D.X., Fan, Y.J., et al., 2021. Holocene moisture evolution and its response to atmospheric circulation recorded by aeolian deposits in the southern Tibetan Plateau. Quat. Sci. Rev. 270, 107169. .
Yang, L., Li, J., Tsamchu, D., 2016. Number and distribution of wintering black-necked crane (Grus nigricollis) in drainage area of Yarlung Zangbo River and its two branches from Tibet, China. J. Northeast For. Univ. 44 (5), 70–72. . (in Chinese).
Yan, R.Q., Shen, W.S., Zhao, W., Ouyang, Y., 2018. Dynamics of suitability and conservation of the black-necked crane habitats in the midstream of the Yarlung Zangbo River. J. Ecol. Rural Environ. 34 (2), 146–152. . (in Chinese).
Yang, Y., Xie, C., Shen, C.Y., Tian, B.S., Wang, S.D., Bian, X.L., et al., 2023. Changes in the landscape patterns of black‐necked crane habitat and its correlation with their individual population numbers during the past 40 years in China. Ecol. Evol. 13, e10125. .
Zhao, Z.L., Liu, L.S., Wang, Z.F., Zhang, Y.L., Li, L.H., Liu, F.G., 2020. Dynamic changes of plateau wetlands in the Damqu River basin, Yangtze River source region, China, 1988-2015. Wetlands 40, 1409–1424.
Zhou, S.L., Liu, S.S., Li, J.R., Yang, L., 2020. Species diversity of wintering waterbirds along Yarlung Zangbo River and its three branches in Tibet. J. Ecol. Rural Environ. 36, 1410–1415. . (in Chinese).
Zhu, L.P., Lü, X.M., Wang, J.B., Peng, P., Kasper, T., Daut, G., et al., 2015. Climate change on the Tibetan Plateau in response to shifting atmospheric circulation since the LGM. Sci. Rep. 5, 13318. .
Table
1.
The number and distribution of Black-necked Cranes in the Yarlung Zangbo River Basin, Tibet, 2018 and 2022.
City
County
Middle wintering period in 2022
Middle wintering period in 2018a
Shigatse
Lhaze
2090
1711
Sakya
212
236
Thongmon
468
308
Samzhubze
1780
2735
Namling
1943
1366
Panam
22
66
Gyantse
0
15
Rinbung
0
1
Subtotal
6515
6438
Lhasa
Quxu
14
17
Taktse
33
574
Lhunzub
1337
830
Medro Gongkar
353
210
Subtotal
1739
1631
Shannan
Gonggar
528
118
Danang
39
2
Nedong
122
63
Sangri
4
39
Subtotal
693
222
Nyingchi
Gongbo Gyamda
86
–
Bayi
10
–
Mainling
296
–
Subtotal
392
–
Total
9337
8291
a Indicates that the data were obtained from Jia et al. (2019); numbers in bold indicates the change of individual numbers between 2018 and 2022 greater than 300; "–" means data not available.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds