Marco Cucco, Giovanni Boano, Carlo Ciani, Mauro Ferri, Luciano Gelfi, Fausto Minelli, Irene Pellegrino. 2025: The impact of climatic conditions in wintering and breeding quarters on long-term breeding success in the Common Swift. Avian Research, 16(1): 100242. DOI: 10.1016/j.avrs.2025.100242
Citation:
Marco Cucco, Giovanni Boano, Carlo Ciani, Mauro Ferri, Luciano Gelfi, Fausto Minelli, Irene Pellegrino. 2025: The impact of climatic conditions in wintering and breeding quarters on long-term breeding success in the Common Swift. Avian Research, 16(1): 100242. DOI: 10.1016/j.avrs.2025.100242
Marco Cucco, Giovanni Boano, Carlo Ciani, Mauro Ferri, Luciano Gelfi, Fausto Minelli, Irene Pellegrino. 2025: The impact of climatic conditions in wintering and breeding quarters on long-term breeding success in the Common Swift. Avian Research, 16(1): 100242. DOI: 10.1016/j.avrs.2025.100242
Citation:
Marco Cucco, Giovanni Boano, Carlo Ciani, Mauro Ferri, Luciano Gelfi, Fausto Minelli, Irene Pellegrino. 2025: The impact of climatic conditions in wintering and breeding quarters on long-term breeding success in the Common Swift. Avian Research, 16(1): 100242. DOI: 10.1016/j.avrs.2025.100242
Climate change can affect rainfall and temperature worldwide, and the ability of birds to react to these changes can be mirrored by studying population phenology and their breeding success. Some European species have advanced arrival and breeding dates in response to local spring advancement, but conditions experienced during winter seasons may also affect arrival dates and subsequent breeding success. We utilized data collected from 1983 to 2020 in four colonies of Common Swift (Apus apus) in Northern Italy (5486 breeding attempts) to examine the variations of laying date and breeding success in relation to non-breeding conditions (previous winter in Africa) and local conditions in breeding period (spring). Climatic conditions were monitored using the North Atlantic Oscillation (NAO) and El Nino Southern Oscillation (ENSO) indices to describe conditions experienced in winter quarters and the local temperature and rainfall conditions of the breeding area. Common Swifts laid their eggs earlier in warmer springs, and this in turn had a positive effect on breeding success. We did not find evidence for any effects of African winter conditions on laying date nor on breeding success. However, because studies made in the same area have demonstrated a relationship of winter conditions (NAO) on individual survival, our data highlight the importance of considering environmental variables across the annual life cycle to understand variation in Common Swift populations.
Climate influences the phenology, reproductive success, population abundance, and geographical distribution of both animal and plant species (Lemoine et al., 2007; Walther, 2010). Due to their high mobility, birds are particularly quick to adapt their phenology, reproduction, and migration strategies in response to climatic changes, such as precipitation and temperature (Romano et al., 2023). The ultimate goal of any such change is to maximize individual reproductive success, a characteristic that shapes the evolutionary process (Alerstam et al., 2003). In boreal areas, Hallfors et al. (2020) found a general advance of avian breeding time, but this pattern was found mainly in resident and short-distance migrating species (Hallfors et al., 2020). For long-distance migratory birds, the optimal timing of the onset of reproduction is vital, especially when suitable conditions for reproduction occur only for a short period during the year. At higher latitudes, the suitable period shortens, leading to greater synchronization of the annual cycle with local conditions among individuals within the same population (Meier et al., 2020).
Usually, production of offspring is timed to coincide with optimal food availability at the time of chick rearing (Lack, 1968). Individuals cannot depend on the arrival of seasonal food availability to begin breeding, but over evolutionary time they have evolved mechanisms to monitor and respond to day length (photoperiod) in order to anticipate seasonal changes in the environment (Walker et al., 2019). Breeding timed to match optimal resource abundance is vital for the successful reproduction of species (Cresswell and McCleery, 2003; Verhulst and Nilsson, 2008), and breeding should therefore be sensitive to environmental cues and should shift in an optimal manner in order to comply with a changing climate (Hallfors et al., 2020). However, adaptations to the photoperiod may be insufficiently plastic to allow a shift in the seasonal timing of bird breeding (Devictor et al., 2008; Walker et al., 2019).
Detailed long-term studies are required to inform of any change of birds’ breeding season. Advancement of the breeding season has been found in several northern hemisphere species (Crick and Sparks, 1999; Cotton, 2003; Potti, 2008, 2009; Hauber et al., 2021). However, this advancement may only be present in some populations of the same species, as demonstrated in an examination of 11 populations of Blue Tit (Cyanistes caeruleus) (Visser et al., 2003; Potti, 2009).
Møller et al. (2006) indicated two hypotheses related to how climate change affects population size: 1) tub-hypothesis, which assumes a strong interference in the non-reproductive period that results in a change in survival in the winter period; 2) tap-hypothesis, which supposes an interference during the reproductive phase which results in a conditioning of the recruitment of new individuals in the following year (Møller et al., 2006). Specifically for the effects of climate, in the tub-hypothesis the variation in population size is associated with the meteorological variation in the non-reproductive season, while in the tap-hypothesis, fluctuations in population size are related to climatic conditions during the reproductive period (Salewski et al., 2013).
In order to relate avian phenology to climatic conditions in different areas of the Earth, suitable climatic variables describing conditions have to be utilized (Remisiewicz and Underhill, 2022). The use of comprehensive climatic indices is spreading because they have the advantage of synthesizing weather conditions into a single variable (Gordo, 2007a). For European swifts, climatic conditions over large parts of the winter quarters have been found to be best described by two climatic indices, the North Atlantic Oscillation (NAO) and the El Nino Southern Oscillation (ENSO) (Åkesson et al., 2020; Boano et al., 2020).
The NAO plays an essential role in the migration of birds throughout the Mediterranean basin, and it is probably a reason for the observed advance of arrival dates during the spring in Europe (Hurrell and Deser, 2009; Gordo et al., 2011). In the breeding period, an influence of NAO has been observed in marine birds, as the availability of food sources such as herring, shellfish, zooplankton, often causes changes in favourable growth and productivity (Abraham and Sydeman, 2004; Bennett et al., 2017). In terrestrial birds, variation in reproduction and breeding related to climate change along the western Palearctic was found in most of the Blue Tit populations, with an advancement of the laying date relative to the winter NAO, although it was more relevant in northern than southern locations (Potti, 2009).
The ENSO represents one of the most significant climatic phenomena of our planet, which strongly influences the regime of winds, temperatures, hydrological state, and the location of high- and low-pressure centers on a planetary level. Two important oceanic components are distinguished: El Niño, associated with warming, and La Niña, associated with cooling (Trenberth, 1997). Duursma et al. (2018) showed a longer laying period during La Niña events, but in other studies no relationships with ENSO (MacMynowski and Root, 2007; Connare and Islam, 2022) or complex relationships including ENSO plus other climatic indices were found (Remisiewicz and Underhill, 2020).
The direct effect of weather conditions during the breeding period (tap-hypothesis) has been reported in several studies (Lack, 1968; Sæther et al., 2004; Barras et al., 2021; Jenouvrier et al., 2022), and appears to be particularly important in aerial feeding birds such as swifts, swallows, and martins (Møller, 1994; Cox et al., 2020; Shipley et al., 2020; Imlay et al., 2022). In the House Martin (Delichon urbica) in Scotland, variables such as temperature, daytime rainfall, and wind intensity described 50% of the daily variation of the flying insects fed to chicks by this aerial feeding bird (Bryant, 1975). In the Common Swift (Apus apus), Lack (1956) showed a strong effect of local poor weather on chick survival. Concerning other swifts’ European species, in the Pallid Swift (Apus pallidus) the frequency of feeding of the chicks was little influenced by the abundance of aerial food because the parents compensated at their own expense in cases of food shortage (Cucco and Malacarne, 1996a, 1996b), while in the Alpine Swift (Tachymarptis melba) the chick’s growth was negatively influenced by adverse weather conditions throughout the development of offspring (Masoero et al., 2024).
No detailed long-term studies are available on the influence of weather conditions on the reproduction of the Common Swift. In this study, we examined the reproduction in four colonies of the Common Swift in Northern Italy, with a long-term perspective using a data set collected up to 38 years. Aim of the research was to examine: ⅰ) if there was a long-term trend to vary the laying date along the study period; ⅱ) if the laying date in each year was related to local weather conditions in the breeding period, or to weather conditions experienced in the winter quarters; ⅲ) if the annual breeding success was related to local weather conditions or to climatic conditions experienced in winter quarters.
2.
Materials and methods
The Common Swift winters below the Equator in Africa (Åkesson et al., 2012). The use of geolocators accurately traced the non-reproductive movements of adults from one of our study colonies, Modena Roccamalatina. The swifts remained in Southeast Africa, and moved between different locations, where they stayed for several days during the winter period (Åkesson et al., 2020; Boano et al., 2020). During the breeding season, the Common Swift is found in a wide area from the Iberian Peninsula to Scandinavia, in countries around the Mediterranean Sea, from North Africa to the Middle East, and in parts of Asia, as far east as China and Siberia. Breeding individuals commonly cluster in colonies, with nests rarely found in natural environments, in cavities in trees or rock cliffs, and much more commonly in the cavities of man-made buildings. Each cavity contains only one nest, because swifts are extremely territorial and defend their cavity from intruders (Cramp, 1985).
Long-term breeding data were collected at four sites in Northern Italy: a) Roccamalatina: the study colony is located in the "Rondonara del Castellaro" tower (Guiglia: 44°23ʹ19ʺ N, 10°57ʹ02ʺ E; Ferri et al., 2014; Minelli et al., 2014), the data include 1371 nests and encompass 29 years in the period 1991–2021, with no sampling in 1997 and 1998; b) Boltiere: the study colony is located in the "Rondonaia" tower (Boltiere: 45°36ʹ03ʺ N, 9°34ʹ51ʺ E), the data include 1005 nests and encompass 15 years in the period from 2006 to 2020; c) Brembate, the study colony is located in the "Berengario" tower (45°36ʹ11ʺ N, 9°33ʹ23ʺ E), the data include 477 nests and encompass 15 years in the period from 2006 to 2020; d) Portico di Romagna, the study colony is located in the "Levante" tower (44°01ʹ32ʺ N, 11°46ʹ54ʺ E; Ciani, 1992), the data include 2633 nests and encompass 38 years in the period from 1983 to 2020. The two breeding colonies in Brembate and Boltiere are 2 km apart, and are located 171 km NW of Roccamalatina, while the colony in Portico is located 78 km SE of Roccamalatina.
The nest cavities were checked from the interior of the buildings with different frequencies in different colonies and years. In most cases, inspections took place several times during the breeding season, from late May to early July. Since the number of eggs or nestlings could vary during the season, each year all four colonies were visited starting during the same week from the beginning of the breeding season until the end. During each inspection all chicks were ringed, and breeding adults were captured and ringed when present. The laying date (Julian date, i.e., days elapsed from January 1st) for each nest was estimated by backdating based on nest content, considering 20 days for egg incubation (Cramp, 1985) and adding the estimated age of the chicks based on growth curves (Tigges, 2008; Sicurella et al., 2015). When the inspections were too far apart to allow backdating of the laying date, we only estimated the fortnight (15-day period) during which laying occurred. Laying fortnights were scheduled as: 0 = 16–30 April; 1 = 1–15 May; 2 = 16–31 May; 3 = 1–15 June; 4 = 16–30 June.
The success of each nest was estimated by counting the number of chicks that were present within each nest at the time of chick ringing. We did not ring very young chicks (naked or with feathers still concealed under the skin). In this case, we returned a few days later, when nestlings were suitable for ringing after their 10th day of life. As mortality of older chicks is usually very low (Dolnik and Kinzhewskaja, 1980; Martins and Wright, 1993a) the estimate of breeding success should be only slightly overestimated.
2.1
Climatic variables
To estimate the effect of climatic conditions on the laying date and the number of chicks reared, we utilized two environmental indices related to the conditions found in the wintering areas, NAO and ENSO, and two environmental indices related to the conditions found in the breeding area, the air temperature and the number of rainy days.
The NAO is a large-scale oscillation in atmospheric masses between the subtropical high and the polar low; the NAO index is based on the difference in winter pressure between a station in Iceland and one in the Azores: positive (negative) values indicate that pressure is higher (lower) than normal over the Azores and lower (higher) than normal over Iceland, representing an increase (decrease) in the North Atlantic Ocean pressure gradient and zonal wind speed. This index is useful as a measure of the general climatic conditions in large parts of Europe (Hurrell et al., 2003). The NAO is considered to also show an influence on rainfall variability (McHugh and Rogers, 2001) and therefore on the crop productivity (Stige et al., 2006) of diverse African regions as a result of atmospheric teleconnections, with positive anomalies associated to drier conditions in north-western and south-eastern Africa, whereas negative anomalies are associated with rainfall surpluses in the same areas. Values of NAO index for the four months from December to March (DJFM) were obtained from https://climatedataguide.ucar.edu/sites/default/files/2023-04/nao_pc_djfm.txt.
The ENSO is a large-scale air-sea variability in the equatorial Pacific. As an index of the El Nino-Southern Oscillation cycle, we used the Oceanic Niño Index (ONI) for the months December, January, and February. ONI is based on sea surface temperature anomalies and is defined as the three-month (in our case Dec-Jan-Feb) running-mean SST (sea surface temperature) departures in the Niño 3.4 region, based on the NOAA Extended Reconstructed Sea Surface Temperature (ERSST) data. El Niño is characterized by ONI ≥ 0.5 ℃, while La Niña is based on ONI ≤ −0.5 ℃. An El Niño or La Niña episode is defined when the above thresholds are exceeded for a period of at least five consecutive overlapping three-month seasons. ENSO is confirmed as having a dominant role in spatial and seasonal rainfall distribution in north-eastern, eastern, and southern Africa (Camberlin et al., 2001). In particular, positive anomalies of ENSO (El Niño conditions) are associated with drier conditions and scarcity of food productions in many parts of Africa from North to South and wetter conditions in eastern Africa, while negative anomalies (La Niña conditions) often brings exceptional rains and exaggerated flooding in Southeast Africa (Stige et al., 2006; Boano et al., 2020). We used ONI data downloaded from https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php.
The climate in the plains of northern part of Italy is temperate, rainfall reaches a maximum in spring and again in autumn. From June to September the weather is generally favourable, with high temperatures and few rainy days (Beck et al., 2018). Local temperature and rainfall were measured along the spring season in Bologna Borgo Panigale Airport (44°31ʹ48ʺ N, 11°17ʹ36ʺ E), near the two study colonies of Roccamalatina and Portico di Romagna (respectively 18 and 68 km distance) and in Bergamo airport (45°40ʹ11ʺ N, 9°42ʹ5ʺ E) near the two study colonies of Boltiere and Brembate (12 and 13 km distance). The mean daily temperature (℃) and the number of rainy days were counted for each nest depending on the precision of the laying date. In the Roccamalatina colony, we utilized the values relative to the precise laying date (N = 1273 cases), while in the other colonies we utilized the values pertaining to the decades when laying occurred (N = 1123 cases). We did not analyze the laying date in relation to weather conditions from the Portico di Romagna colony because the inspection schedule did not all allow a precise estimation of the laying date.
2.2
Statistics
First, we examined the relationship between the dependent variables (laying date and breeding success) and the climatic variables during the preceding winter period (ENSO and NAO) or the local weather variables by means of single-variable regressions. As performing many statistical tests increases the probability of obtaining false positives (type Ⅰ error), we applied Bonferroni correction (Rice, 1989) to the P-values. Secondly, we utilized general linear models to examine the effect of temperature, rainfall, ENSO and NAO indices on laying date and breeding success. Initially, we entered all climatic variables simultaneously. However, temperature or rainfall values in different fortnights can be highly correlated. Hence, we used a stepwise backward procedure of selection of independent variables by eliminating, at each step, the variables that had the smallest predictive power. The stepwise backward procedure led to a final model containing only the variables reaching the 0.05 level of significance. Statistics were calculated in the R software environment (R Core Team, 2023) with the lme4 package (Bates et al., 2015).
To test for significant difference between colonies in the number of females laying in different periods, we utilized the chi-square statistic.
Differences of annual mean number of chicks in the different colonies were tested with ANOVA, and multiple comparisons of means between colonies were conducted by Tukey contrasts in the R software package multcomp (Hothorn et al., 2008).
3.
Results
3.1
Laying date
Common Swifts laid their eggs mainly in May (Table 1). We found a significant difference in laying date between colonies (Chi-square test = 170.8, d.f. = 8, P < 0.001), with most females of Roccamalatina laying earlier that those in Boltiere and Brembate.
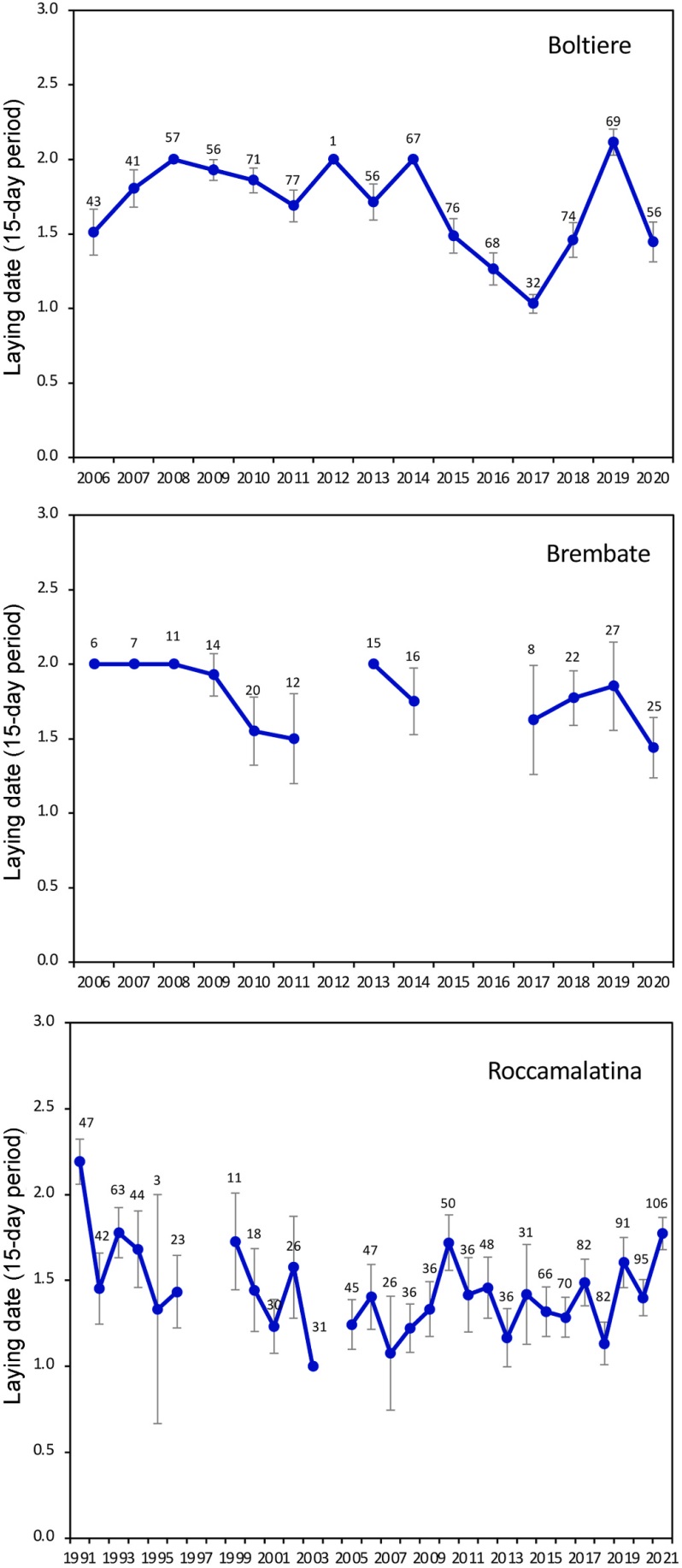
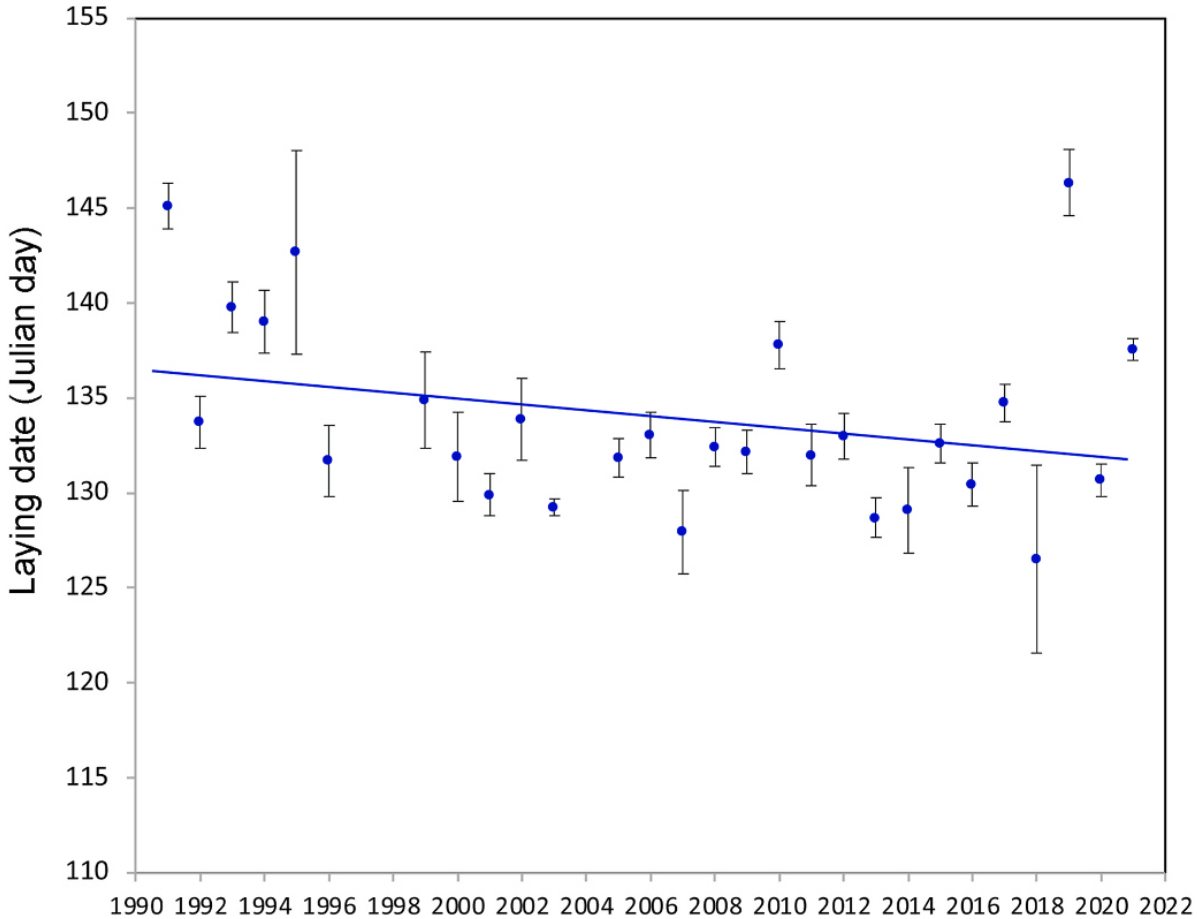
We found significant differences in laying date (fortnights) among years (Fig. 1; Roccamalatina: F27,1293 = 9.22, P < 0.001; Boltiere: F14,834 = 31.02, P < 0.001; Brembate: F11,171 = 2.88, P < 0.0017). A significant interannual difference in laying dates (Julian days) was also found considering the more detailed data collected in Roccamalatina (Fig. 2; F27,1105 = 11.56, P < 0.001). The mean laying date in Roccamalatina was the 134th Julian day, i.e., May 15, with a significant tendency to advance 0.12599 ± 0.02892 day/year (F1,1131 = 4.36, P < 0.001), with a total estimated advancement of 3.906 days over the 31-year study period.
Figure
1.
Laying period (fortnights, 15-day periods) in different years spanning from 1991 to 2021. Code of the 15-days periods: 0: 15–30 April; 1: 1–15 May; 2: 16–31 May; 3: 1–15 June. Numbers over the dots indicate the number of sampled nests, bars represent the 95% confidence limits.
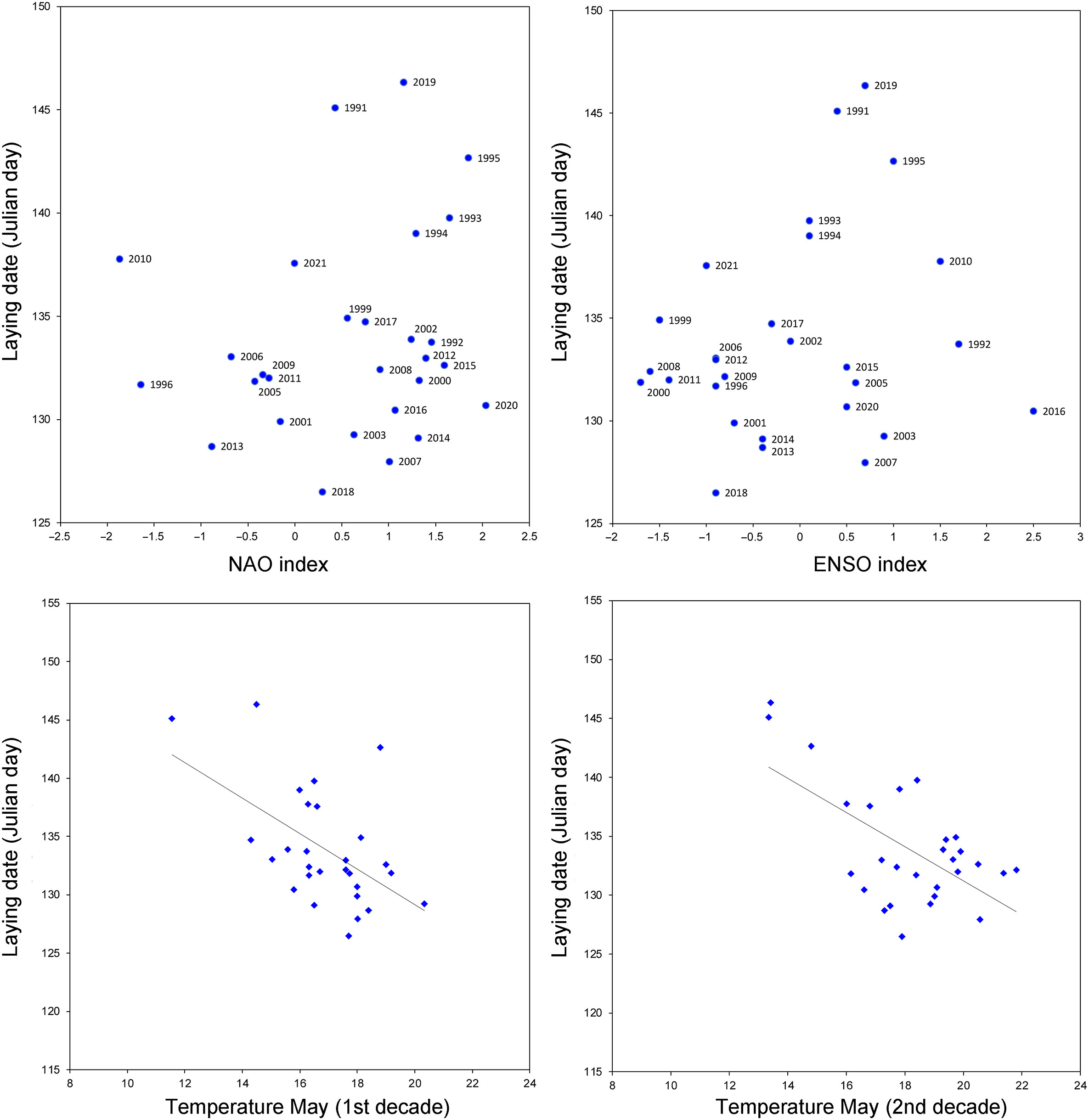
The climatic conditions experienced during winter in the African continent did not significantly influence the mean laying date (Julian day) of the Common Swift in Roccamalatina (Fig. 3). This was observed both for the NAO and the ENSO indices (Table 2).
Figure
3.
Relationship between mean laying date in different years at Roccamalatina and climatic indices during the winter (NAO and ENSO) and temperature and rainfall during the reproductive period.
Table
2.
Relationship between mean annual laying date (Julian day) in Roccamalatina and NAO or ENSO indices in the winter period, and local temperature or local rainfall during selected decades.
The temperatures of the third decade of May and the first decade of June did not significantly affect the mean annual laying date (Table 2). On the contrary, the temperatures of late April and early May, right in the full laying period, were negatively related with the mean laying date (Fig. 3). In years in which the recorded temperatures were low, the dates had higher values, i.e., they occurred later, with an average delay of 1.41–1.57 days for every one degree decrease in temperature (Table 2).
Local rainfall was not significantly related with the mean annual laying date, with the exception of the second decade of May (single variable model), during which high rainfall values were related to later laying dates (Table 2). In the multivariate model, constructed by including all winter climate variables (NAO and ENSO) and breeding period weather variables (temperature and rainfall), the stepwise procedure only retains the temperature of the first and second decade of May with statistically significant values (Table 2).
3.2
Breeding success
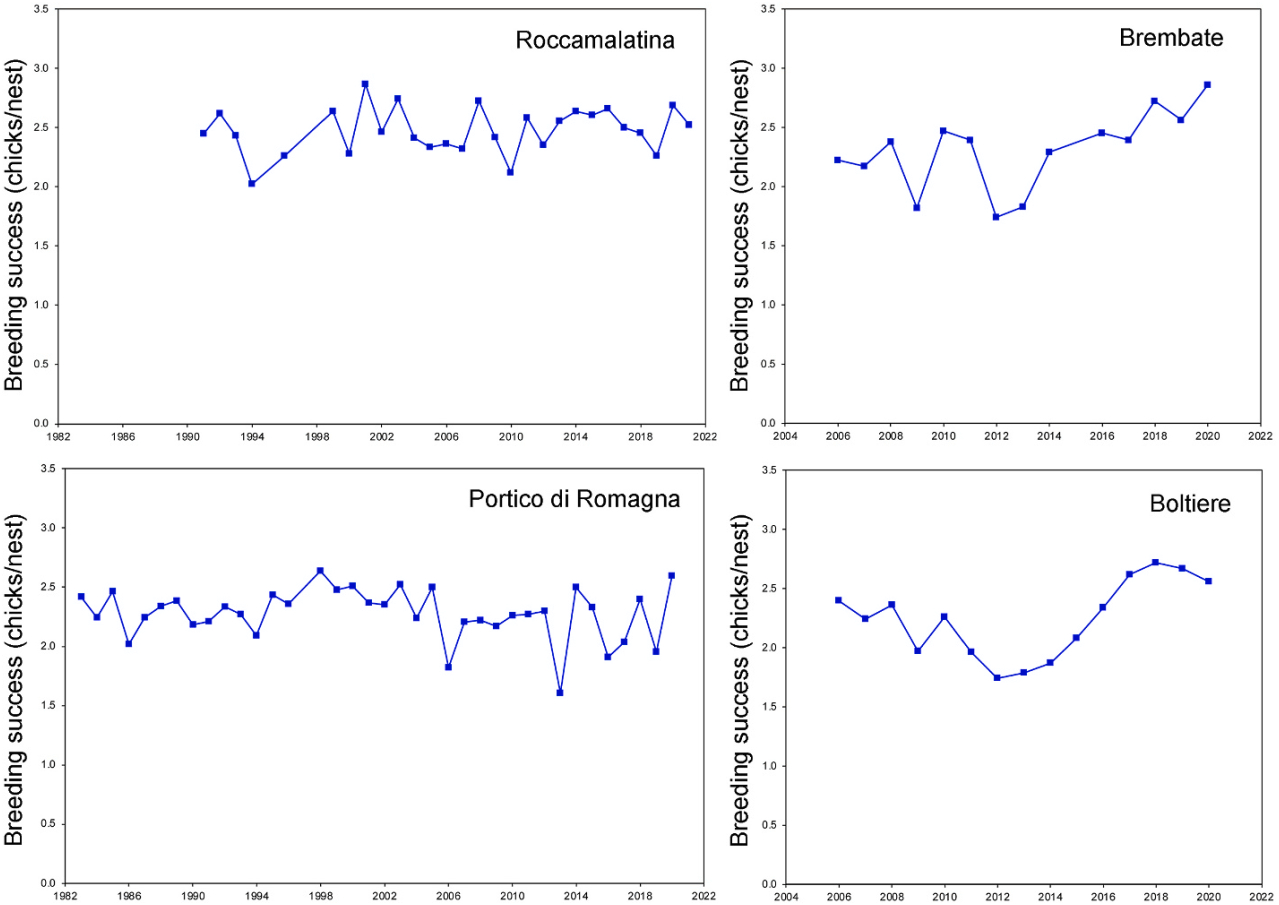
The mean number of chicks per nest was 2.474 ± 0.194 in Roccamalatina, 2.239 ± 0.325 in Boltiere, 2.306 ± 0.330 in Brembate, and 2.275 ± 0.218 in Portico di Romagna (Fig. 4). The four colonies differ in the number of chicks/nest both when comparing all the years available in the different colonies (F3,90 = 4.385, P = 0.0063) and when comparing the 14 years that were sampled concurrently in all four colonies (F3,52 = 2.994, P = 0.039). A multiple comparison of means showed that a significant difference was found between Roccamalatina and Portico (Tukey contrasts: estimate −0.199 ± 0.063, t = −3.174, P = 0.011) and between Roccamalatina and Boltiere (Tukey contrasts: estimate −0.235 ± 0.080, t = −2.937, P = 0.02), while all other contrasts between colonies showed intermediate values and did not differ significantly (Tukey contrasts: all P > 0.05 n.s.).
Figure
4.
Breeding success (chicks/nest) in different years spanning from 1991 to 2021 in four colonies in Northern Italy.
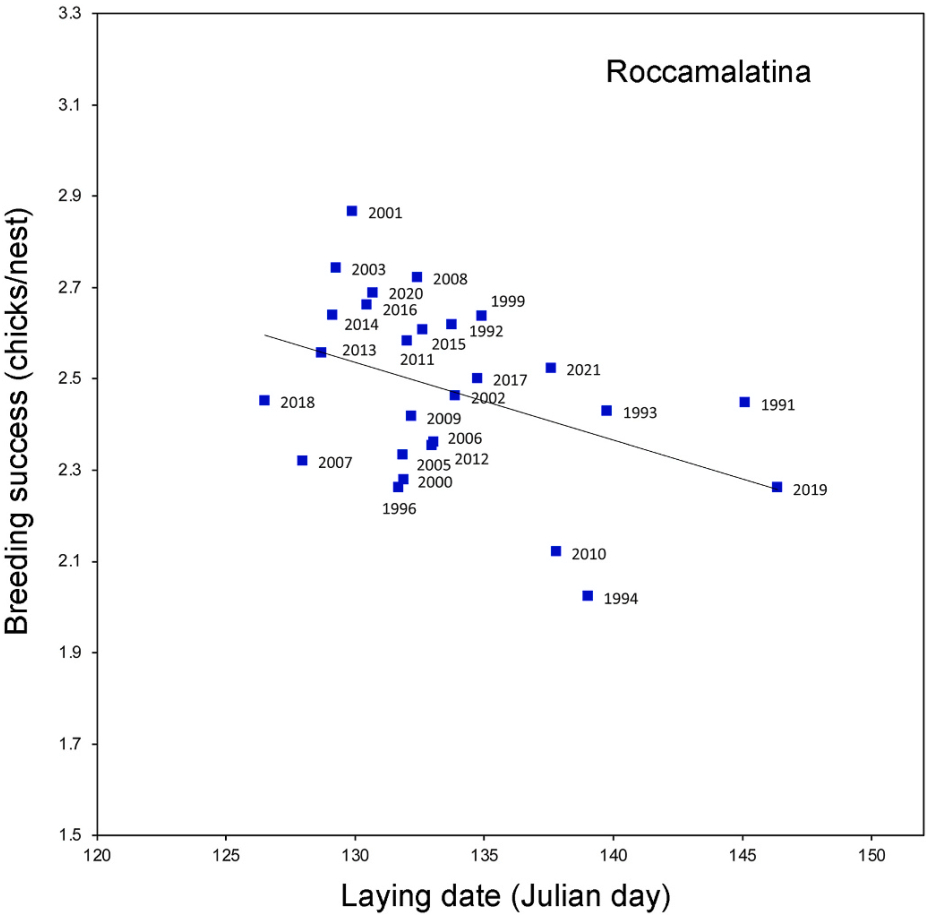
In the Roccamalatina colony, there was a negative correlation between mean laying date and reproductive success (F1,25 = 5.088, P = 0.033) with smaller brood sizes when a delay of laying occurred (Fig. 5).
Figure
5.
Relationship between mean laying date and breeding success at Roccamalatina colony.
The breeding success in the four study colonies was unrelated to climatic conditions experienced during the winter period in Africa (Appendix Table S1). Weather conditions experienced in spring during the breeding period (from late April to June) show a positive relationship between breeding success and temperature in the first decade of June at Portico di Romagna colony (Appendix Table S1). In the multivariate model, constructed by including all the winter climate variables (NAO and ENSO) and the weather variables of the breeding period (temperature and rainfall), the stepwise procedure only retains the temperature of the first decade of June with statistically significant values in the colony of Portico di Romagna (Appendix Table S1), while there was no relationship between weather variables and breeding success in the other colonies.
4.
Discussion
Our study shows that climatic conditions experienced by Common Swifts during the non-breeding period in Africa did not influence the subsequent breeding season, in terms of laying date nor as the number of chicks reared. Low temperature conditions experienced during the spring breeding period could postpone the laying date, with negative effects on the breeding success.
The lack of relationship between wintering climate and reproductive parameters could suggest that our data is in line with the tap-hypothesis (Møller et al., 2006), which predicts that the variation in population size is associated with the meteorological variation in the reproductive season, while in the tup-hypothesis, fluctuations are related to climatic conditions during the non-reproductive period (Salewski et al., 2013). However, when modelling population dynamics, the whole life cycle must be considered, and the conditions that influence breeding success must be flanked with those influencing the survival of individuals (McGraw and Caswell, 1996; Knudsen et al., 2011). In our study species, swift survival varies between years and is strongly affected by climatic drivers (ENSO) associated with its wintering area (Boano et al., 2020). Therefore, when considering the total annual cycle, the climatic conditions in the non-breeding season seem to act with a large influence on the Common Swift life history.
In this study, we observed a long-term trend toward advancing laying date across years. This result is in line with several studies showing that birds tend to lay eggs earlier (Crick et al., 1997; Cotton, 2003; Gordo, 2007a). Rapid climate change has been implicated as a cause of modifications in bird population schedules. Some long-term studies, spanning over several decades on a single species, have shown that populations are able to track a rapidly changing environment very closely (Crick et al., 1997; Both et al., 2004; Dunn, 2004; Gordo, 2007b; Charmantier et al., 2009).
In comparison to laying dates, many more studies analysed the temporal trend in arrival date (Møller et al., 2008). This is a parameter easier to collect in the field with respect to laying date, because one can benefit from data gathered through simple observations of bird first annual presence without searching for nests and inspecting their content to assess egg occurrence. Migratory phenology shifted during the last decades (Sparks, 1999) in response to the increase of spring temperature, the advancement of food availability, and the plant phenology in the breeding grounds (Menzel and Fabian, 1999). An early arrival can benefit individuals in terms of territory and mate acquisition (Møller, 1994; Kokko, 1999). However, an early arrival may not translate directly into early laying date. The production of offspring must be timed to coincide with optimal food availability, and physiological mechanisms should evolve in order to anticipate seasonal changes in the environment, e.g., responding to reliable indices of season advancement, such as day length (Walker et al., 2019). Indeed, over evolutionary time, there has been precise fine-tuning of critical photoperiod and onset/offset of seasonal adaptations.
As regards the European Apodidae, the data recorded from the 1970s onwards for the Pallid Swift in NW Italy has verified a continuous advancement of the first arrivals, moving from the first ten days of April to the last ten days of March, with an advancement of about four days per decade (Boano and Perosino, 2014). The arrival of the Common Swift from the African wintering territories appears to be staggered according to latitude. In Southern Italy, the migratory arrival is around the second part of March, while in Northern Italy, it occurs in the month of April (Boano and Cucco, 1989; Brichetti and Caffi, 1994), with arrival advancing by about three days per decade in the period 1986–2006 (Rubolini et al., 2007). In Spain the advancement of dates, in many cases, has shown a strong negative correlation with local temperature, as well as with the rainfall in the Sahel during the previous season (Gordo and Sanz, 2006). However, other long-term observations in several European cities had not recorded a change in the arrival dates, but instead periodic shifts in terms of anticipation and postponement every 15–20 years (Gordo, 2007b). In the Alpine Swift, arrival and length of stay at the breeding sites was negatively correlated with latitude and differed by more than a month between populations (Meier et al., 2020).
A consequence of phenological anticipation in arrivals for reproduction has been a common trend towards an advance of autumn departures for African wintering areas (Jenni and Kéry, 2003). The advancement leads to early reproduction and consequently to an early departure to make the most of the favourable environmental conditions in Africa (Jenni and Kéry, 2003; Gordo and Sanz, 2006).
After the arrival from the winter quarters, local weather can influence the start of egg laying (Lack, 1968; Visser and Both, 2005). In several species, the general advancement of season was accompanied by an advancement of breeding date, but several cases of temporally decoupled weather metrics and bird reproductive onsets (Hauber et al., 2021) or lack of advancement were reported (Gordo, 2007b).
In our study, the temperatures of late April and early May, right in the full reproduction period, were significantly related with the laying date of Common Swifts. In years in which the recorded temperatures were low, the egg laying occurred later, and vice versa. Local rainfall was not found to be significantly related with the laying date, probably because rainwater has a double contrasting effect on environmental conditions for swifts at our study site latitude, reducing the flying invertebrates that are food for these aerial feeders during the rainfall hours, but increasing the growth of vegetation and the number of insects which consume vegetal substances after rainfall events (Cocu et al., 2005; Bourhis et al., 2021).
The breeding success in our study area was similar to those reported in other portions of the breeding range (Lack, 1956; Weitnauer, 1980; Cramp, 1985; Meyer, 2019; Knaus et al., 2022), with small differences between colonies. In our long-term study, we did not notice a decline in the breeding success, similarly to a long-term study in Scotland (Thomson et al., 1996), while in Bohemia, a decline of productivity was found (Rajchard et al., 2006). The reproductive success was significantly influenced by the laying date, with the number of chicks per nest decreasing when a delay occurred. Higher temperatures in the laying period were related to a double positive effect, due to a predisposition to advance laying, and this in turn was matched with an increase in fitness. Rainfall was not negatively related to breeding success, contrary to that reported in northern areas (Koskimies, 1950; Lack and Lack, 1951; Lack, 1956; O’Connor, 1979) and in Bohemia (Rajchard et al., 2006). This is probably because rain can be beneficial to vegetal biomass and production of invertebrate aerial plankton in regions where climate during the breeding season is typically hot and characterized by low rainfall (Sicurella et al., 2015), while long periods of bad weather conditions that can lend to starvation (Gory, 1987; Thomson et al., 1996) are less likely to occur.
Our result is similar to that recorded in the Pallid Swift in the same geographical area, where laying dates were influenced by the weather in terms of unfavourable temperatures recorded in May, causing a delay of laying until June, as well as by the presence of rainfall, with an optimum number of 8–10 rainy days in the period of chick rearing, while years with scarce or excessive rainfall lend to a lower breeding success (Cucco et al., 1992).
We did not find any influence of climatic conditions during the winter stay in Africa on successive breeding success. This contrasts with findings on some passerines (Ockendon et al., 2014) but highlights the necessity to include several drivers both in Africa and in Europe to understand the populational variation of birds (Millon et al., 2019; Howard et al., 2020).
A complete analysis of the influence of climatic conditions on the fitness of Common Swifts should consider other life-history traits, such as survival, in addition to annual breeding success. Martins and Wright (1993b) showed that, in poor weather, the food was limited and that parents allocated food to their brood and sacrificed their own self-feeding. The percentage of swifts in the diet of predatory birds can increase by more than 10-fold at the end of breeding seasons or after harsh weather (Kuhk, 1948; Klass, 1953). In Scotland, a simulation integrating the effects of conditions around the study site, breeding success and annual survival, suggests that lifetime reproductive success is influenced more by variations in lifespan than by annual breeding success (Thomson et al., 1996).
Survival of the Common Swift is also negatively influenced by conditions in the winter quarters, as summarized by ENSO index (Boano et al., 2020). Furthermore, winter survival is strongly dependent on particularly extreme conditions, such as heavy rainfall in the African regions where the species is present during this period (Boano et al., 2020).
The importance of climatic conditions in the precise areas crossed by each species is highlighted by the findings on the Pallid Swift, in which African wintering areas differ from the Common Swift and whose annual survival is linked to other summarizing indices (the precipitation of the Sahel and the NAO: Boano et al., 2020).
Recently, a possible influence of conditions during the migratory itinerary has been highlighted on birds (Taylor et al., 2016). However, no studies have examined the effects of weather during migration on the swifts, even if the remarkably fast crossing of desert and other unfavourable areas suggests a less pronounced effect (Åkesson and Bianco, 2021; Finlayson et al., 2021).
5.
Conclusions
The laying date of our study species was influenced by the long-term increase of temperatures in the breeding area, showing a tendency towards anticipation of about four days along the study period. Reproductive success was significantly influenced by the laying date, which in turn was annually influenced by local temperatures. Recently, Hadfield and Reed (2022) point out that a mismatch between when individuals breed and when they should breed can be a problem in evolutionary ecology. Directional selection can move the optimum across years (Moiron et al., 2024), but when there is large annual variation in climatic conditions then phenological plasticity can evolve to cope with fluctuations in the optimum (de Villemereuil et al., 2020). A genetic selection to advance the laying date with a warming spring may get breeding parents to face a greater risk of chick mortality later (Shipley et al., 2020), but this apparently will not apply to the case of Common Swift in North Italy, because current projections of future climate in this part of Europe predict an intensification of drought periods (Baronetti et al., 2022), with negative effects on individuals which will not advance their breeding season.
CRediT authorship contribution statement
Marco Cucco: Writing – original draft, Supervision, Project administration, Methodology, Data curation, Conceptualization. Giovanni Boano: Supervision, Methodology, Investigation, Conceptualization. Carlo Ciani: Investigation. Mauro Ferri: Supervision, Investigation. Luciano Gelfi: Investigation. Fausto Minelli: Investigation. Irene Pellegrino: Writing – original draft, Investigation.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank Giuseppe Rossi, Stefano Sirotti, Mauro Villani, and Romano Benassi for ringing activity in the Roccamalatina colony, and Dr. Franco Colnago, Walter Frigeni, Giancarlo Leidi, Osvaldo Torri in the Brembate and Boltiere colonies. We also would thank all other volunteers helping with the ringing activity at the colonies, and Lisa Berta and Simone Mulè for help in data filing.
Abraham, C.L., Sydeman, W.J., 2004. Ocean climate, euphausiids and auklet nesting: inter-annual trends and variation in phenology, diet and growth of a planktivorous seabird, Ptychoramphus aleuticus. Mar. Ecol. Prog. Ser. 274, 235–250. .
Åkesson, S., Klaassen, R., Holmgren, J., Fox, J.W., Hedenström, A., 2012. Migration routes and strategies in a highly aerial migrant, the Common Swift Apus apus, revealed by light-level geolocators. PLoS One 7, e41195. .
Åkesson, S., Atkinson, P.W., Bermejo, A., de la Puente, J., Ferri, M., Hewson, C.M., et al., 2020. Evolution of chain migration in an aerial insectivorous bird, the common swift Apus apus. Evolution 74, 2377–2391. .
Alerstam, T., Hedenstrom, A., Åkesson, S., 2003. Long-distance migration: evolution and determinants. Oikos 103, 247–260. .
Baronetti, A., Dubreuil, V., Provenzale, A., Fratianni, S., 2022. Future droughts in northern Italy: high-resolution projections using EURO-CORDEX and MED-CORDEX ensembles. Climat. Chang. 172, 1–22. .
Barras, A.G., Niffenegger, C.A., Candolfi, I., Hunziker, Y.A., Arlettaz, R., 2021. Nestling diet and parental food provisioning in a declining mountain passerine reveal high sensitivity to climate change. J. Avian Biol. 52, e02649. .
Bates, D., Mächler, M., Bolker, B., Walker, S., 2015. Fitting linear mixed-effects models using lme4. J. Stat. Software 67, 1–48. .
Beck, H., Zimmermann, N., McVicar, T., Vergopolan, N., Berg, A., Wood, E.F., 2018. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 5, 180214. .
Bennett, J.L., Jamieson, E.G., Ronconi, R.A., Wong, S.N.P., 2017. Variability in egg size and population declines of Herring Gulls in relation to fisheries and climate conditions. Avian Conserv. Ecol. 12, 16. .
Boano, G., Cucco, M., 1989. Breeding biology of the pallid swift (Apus pallidus) in North-western Italy. Gerfaut 79, 133–148.
Boano, G., Pellegrino, I., Ferri, M., Cucco, M., Minelli, F., Åkesson, S., 2020. Climate anomalies affect annual survival rates of swifts wintering in sub-Saharan Africa. Ecol. Evol. 10, 7916–7928. .
Boano, G., Perosino, G.C., 2014. Cambiamenti locali del clima e date di arrivo del rondone pallido (Apus pallidus) in Piemonte. Riv. Piemontese Storia Nat. 35, 269–284.
Both, C., Artemyev, A.V., Blaauw, B., Cowie, R.J., Dekhuijzen, A.J., Eeva, T., et al., 2004. Large-scale geographical variation confirms that climate change causes birds to lay earlier. Proc. R. Soc. B 271, 1657–1662.
Bourhis, Y., Bell, J.R., van den Bosch, F., Milne, A.E., 2021. Artificial neural networks for monitoring network optimisation-a practical example using a national insect survey. Environ. Model. Software 135, 104925. .
Brichetti, P., Caffi, M., 1994. Biologia riproduttiva di una popolazione di Rondone, Apus apus, nidificante in una piccionaia della pianura lombarda. Riv. Italiana Ornitol. 64, 21–27.
Bryant, D.M., 1975. Breeding biology of House martins Delichon urbica in relation to aerial insect abundance. Ibis 117, 180–216. .
Camberlin, P., Janicot, S., Poccard, I., 2001. Seasonality and atmospheric dynamics of the teleconnection between African rainfall and tropical sea-surface temperature: Atlantic vs. ENSO. Int. J. Climatol. 21, 973–1005. .
Charmantier, A., McCleery, R.H., Cole, L.R., Perrins, C., Kruuk, L.E.B., Sheldon, B.C., 2009. Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 320, 800–803. .
Ciani, C., 1992. Dati sulla biologia riproduttiva del Rondone, Apus apus, in Romagna. Rivista Italiana di Ornitologia 62, 171–177.
Cocu, N., Harrington, R., Rounsevell, M.D.A., Worner, S.P., Hulle, M., 2005. Geographical location, climate and land use influences on the phenology and numbers of the aphid, Myzus persicae, in Europe. J. Biogeogr. 32, 615–632. .
Connare, B.M., Islam, K., 2022. Failure to advance migratory phenology in response to climate change may pose a significant threat to a declining Nearctic-Neotropical songbird. Int. J. Biometeorol. 66, 803–815. .
Cotton, P.A., 2003. Avian migration phenology and global climate change. Proc. Nat. Acad. Sci. USA 100, 12219–12222. .
Cox, A.R., Robertson, R.J., Rendell, W.B., Bonier, F., 2020. Population decline in tree swallows (Tachycineta bicolor) linked to climate change and inclement weather on the breeding ground. Oecologia 192, 713–722. .
Cramp, S., 1985. The Birds of the Western Palearctic, vol. 4. Oxford University Press, Oxford.
Crick, H.Q.P., Sparks, T.H., 1999. Climate change related to egg-laying trends. Nature 399, 423–424.
Cresswell, W., McCleery, R., 2003. How great tits maintain synchronization of their hatch date with food supply in response to long-term variability in temperature. J. Anim. Ecol. 72, 356–366.
Crick, H.Q.P., Dudley, C., Glue, D.E., Thomson, D.L., 1997. UK birds are laying eggs earlier. Nature 388, 527, 527.
Cucco, M., Malacarne, G., 1996a. Reproduction of the pallid swift (Apus pallidus) in relation to weather and aerial insect abundance. Ital. J. Zool. 63, 247–253.
Cucco, M., Malacarne, G., 1996b. Effect of food availability on nestling growth and fledging success in manipulated pallid swift broods. J. Zool. 240, 141–151.
Cucco, M., Malacarne, G., Orecchia, G., Boano, G., 1992. Influence of weather conditions on Pallid Swift Apus pallidus breeding success. Ecography 15, 184–189.
Devictor, V., Julliard, R., Couvet, D., Jiguet, F., 2008. Birds are tracking climate warming, but not fast enough. Proc. R. Soc. B 275, 2743–2748. .
de Villemereuil, P., Charmantier, A., Arlt, D., Bize, P., Brekke, P., Brouwer, L., et al., 2020. Fluctuating optimum and temporally variable selection on breeding date, in birds and mammals. Proc. Nat. Acad. Sci. USA 117, 31969–31978. .
Dolnik, V.R., Kinzhewskaja, L.I., 1980. Time and energy budgets during the nest period in the swift (Apus apus) and swallows (Delichon urbica, Hirundo rustica). Zool. Zhurnal 59, 1841–1851.
Duursma, D.E., Gallagher, R.V., Griffith, S.C., 2018. Effects of El Nino southern oscillation on avian breeding phenology. Divers. Distrib. 24, 1061–1071. .
Ferri, M., Minelli, F., Villani, M., Sirotti, S., Rossi, G., Benassi, R., et al., 2014. Variazioni nel numero di nidiate di rondone comune Apus apus in una rondonara storica, nel parco regionale dei sassi di Roccamalatina (Guiglia, MO), nel periodo 1991-2011. In: Tinarelli, R., Andreotti, A., Baccetti, N., Melega, L., Roscelli, F., Serra, L., Zenatello, M. (Eds.), Atti XVI Convegno Italiano di Ornitologia. Cervia (RA), 22-25 settembre 2011. Scritti, Studi e Ricerche di Storia Naturale della Repubblica di San Marino, pp. 360–361.
Finlayson, S., Holmes, T.L., Finlayson, G., Guillem, R., Perez, C., Bensusan, K., et al., 2021. Birds with multiple homes. The annual cycle of the pallid swift (Apus pallidus brehmorum). PLoS One 16, e0259656. .
Gordo, O., 2007a. Does climate change effect Common Swift Apus apus migratory phenology? APUSlife 3237, 1–13.
Gordo, O., 2007b. Why are bird migration dates shifting? A review of weather and climate effects on avian migratory phenology. Clim. Res. 35, 37–58. .
Gordo, O., Barriocanal, C., Robson, D., 2011. Ecological impacts of the North Atlantic oscillation (NAO) in mediterranean ecosystems. In: Vicente-Serrano, S., Trigo, R. (Eds.), Hydrological, Socioeconomic and Ecological Impacts of the North Atlantic Oscillation in the Mediterranean Region, Advances in Global Change Research, vol. 46. Springer, Dordrecht, pp. 153–170. .
Gordo, O., Sanz, J.J., 2006. Climate change and bird phenology: a long-term in the Iberian Peninsula. Glob. Change Biol. 12, 1993–2004.
Gory, G., 1987. Influence du climat méditerranéen sur la reproduction du martinet noir (Apus apus L.). L’Oiseau et la Revue Francaise D’Ornithol 57, 69–84.
Hadfield, J.D., Reed, T.E., 2022. Directional selection and the evolution of breeding date in birds, revisited: hard selection and the evolution of plasticity. Evol. Lett. 6, 178–188. .
Hallfors, M.H., Antao, L.H., Itter, M., Lehikoinen, A., Lindholm, T., Roslin, T., et al., 2020. Shifts in timing and duration of breeding for 73 boreal bird species over four decades. Proc. Nat. Acad. Sci. USA 117, 18557–18565. .
Hauber, M.E., Elek, Z., Moskát, C., 2021. Advancing onset of breeding dates in brood parasitic common cuckoos and their great reed warbler hosts over a 22-year period. Ethol. Ecol. Evol. 33, 553–560. .
Hothorn, T., Bretz, F., Westfall, P., 2008. Simultaneous inference in general parametric models. Biom. J. 50, 346–363. .
Howard, C., Stephens, P.A., Pearce-Higgins, J.W., Gregory, R.D., Butchart, S.H.M., Willis, S.G., 2020. Disentangling the relative roles of climate and land cover change in driving the long-term population trends of European migratory birds. Divers. Distrib. 26, 1442–1455. .
Hurrell, J.W., Deser, C., 2009. North Atlantic climate variability: the role of the North Atlantic oscillation. J. Mar. Syst. 78, 28–41. .
Hurrell, J.W., Kushnir, Y., Ottersen, G., Visbeck, M., 2003. An overview of the North Atlantic oscillation. In: Hurrell, J.W., Kushnir, Y., Ottersen, G., Visbeck, M. (Eds.), The North Atlantic Oscillation: Climatic Significance and Environmental Impact, vol. 134. Geophysical Monograph Series, American Geophysical Union, pp. 1–36.
Imlay, T.L., Mann, H.A.R., Ding, A.C., Thomas, P., Whittam, R.M., Leonard, M.L., et al., 2022. Annual adult survival rates for four sympatric breeding swallow species: effects of environmental factors and density-dependence. Can. J. Zool. 100, 647–659. .
Jenni, L., Kéry, M., 2003. Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proc. R. Soc. B 270, 1467–1471. .
Jenouvrier, S., Long, M.C., Coste, C.F.D., Holland, M., Gamelon, M., Yoccoz, N.G., et al., 2022. Detecting climate signals in populations across life histories. Glob. Change Biol. 28, 2236–2258. .
Klass, C., 1953. Zur Ernahrung des Baumfalken. Vogelwelt 74, 48–49.
Knaus, P., Sattler, T., Schmid, H., Strebel, N., Volet, B., 2022. The State of Birds in Switzerland: Report 2022. Swiss Ornithological Institute, Sempach.
Knudsen, E., Lindén, A., Both, C., Jonzén, N., Pulido, F., Saino, N., et al., 2011. Challenging claims in the study of migratory birds and climate change. Biol. Rev. 86, 928–946. .
Kokko, H., 1999. Competition for early arrival in migratory birds. J. Anim. Ecol. 68, 940–950. .
Koskimies, J., 1950. The life of the swift, Micropus apus (L.), in relation to the weather. Ann. Acad. Sci. Fenn. 15, 1–151.
Kuhk, R., 1948. Wirkungder Regen-und Kalteperiode 1948 auf den Mauersegler, Micropus apus (L.). Vogelwarte 10, 28–30.
Lack, D., 1956. Swifts in a Tower. Chapman & Hall, London.
Lack, D., 1968. Ecological Adaptations for Breeding in Birds. Methuen and Co, London.
Lack, D., Lack, E., 1951. The breeding biology of the Swift Apus apus. Ibis 93, 501–546.
Lemoine, N., Schaefer, H.C., Bohning-Gaese, K., 2007. Species richness of migratory birds is influenced by global climate change. Global Ecol. Biogeogr. 16, 55–64. .
Martins, T.L.F., Wright, J., 1993a. Cost of reproduction and allocation of food between parent and young in the swift (Apus apus). Behav. Ecol. 4, 213–223. .
Martins, T.L.F., Wright, J., 1993b. Brood reduction in response to manipulated brood sizes in the common swift (Apus apus). Behav. Ecol. Sociobiol. 32, 61–70. .
Masoero, G., Dumas, M.N., Martin, J.G.A., Bize, P., 2024. Trait-specific sensitive developmental windows: wing growth best integrates weather conditions encountered throughout the development of nestling Alpine swifts. Ecol. Evol. 14, e11491. .
McGraw, J.B., Caswell, H., 1996. Estimation of individual fitness from life-history data. Am. Nat. 147, 47–64.
McHugh, M.J., Rogers, J.C., 2001. North Atlantic oscillation influence on precipitation variability around the Southeast African convergence zone. J. Clim. 14, 3631–3642.
MacMynowski, D.P., Root, T.L., 2007. Climate and the complexity of migratory phenology: sexes, migratory distance, and arrival distributions. Int. J. Biom. 51, 361–373.
Meier, C.M., Karaardic, H., Aymi, R., Peev, S.G., Vitvliet, W., Liechti, F., 2020. Population-specific adjustment of the annual cycle in a super-swift trans-Saharan migrant. J. Avian Biol. 51, e02515. .
Menzel, A., Fabian, P., 1999. Growing season extended in Europe. Nature 397, 659.
Meyer, S., 2019. Ansiedlungsverhalten des Mauerseglers Apus apus in der Kirche Oberkirch in der Nordwestschweiz. Ornithol. Beob. 116, 1–10.
Millon, A., Danovaro, C., Printemps, T., Leroux, M.B., Schlain, A.E., Villers, A., et al., 2019. Disentangling the effects of environmental conditions on wintering and breeding grounds on age-specific survival rates in a trans-Saharan migratory raptor. J. Avian Biol. 50, e02233. .
Minelli, F., Ferri, M., Villani, M., Sirotti, S., Rossi, G., Benassi, R.F., 2014. Attività di inanellamento in colonie di rondone comune Apus apus in rondonare artificiali storiche nell’area del Parco Regionale dei Sassi di Roccamalatina (Modena), 1991–2011. Scritti, Studi e Ricerche di Storia Naturale della Repubblica di San Marino, 399–400.
Moiron, M., Teplitsky, C., Haest, B., Charmantier, A., Bouwhuis, S., 2024. Micro-evolutionary response of spring migration timing in a wild seabird. Evol. Lett. 8, 8–17. .
Møller, A.P., 1994. Phenotype-dependent arrival time and its consequences in a migratory bird. Behav. Ecol. Sociobiol. 35, 115–122. .
Møller, A.P., Fielder, W., Berthold, P., 2006. Birds and Climate Change. Academic Press, London.
Møller, A.P., Merilä, J., 2004. Analysis and interpretation of long-term studies investigating responses to climate change. Adv. Ecol. Res. 35, 111–130.
Møller, A.P., Rubolini, D., Lehikoinen, E., 2008. Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc. Nat. Acad. Sci. USA 105, 16195–16200. .
Ockendon, N., Johnston, A., Baillie, S.R., 2014. Rainfall on wintering grounds affects population change in many species of Afro-Palaearctic migrants. J. Ornithol. 155, 905–917.
O’Connor, R.J., 1979. Egg weights and brood reduction in the European Swift (Apus apus). Condor 81, 133–145.
Palm, V., Leito, A., Truu, J., Tomingas, O., 2009. The spring timing of arrival of migratory birds: dependence on climate variables and migration route. Ornis Fenn. 86, 97–108.
Potti, J., 2008. Temperature during egg formation and the effect of climate warming on egg size in a small songbird. Acta Oecol. 33, 387–393.
Potti, J., 2009. Advanced breeding dates in relation to recent climate warming in a Mediterranean montane population of Blue Tit Cyanistes caeruleus. J. Ornithol. 150, 893–901.
R Core Team, 2023. R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Rajchard, J., Procházka, J., Kindlmann, P., 2006. Long-term decline in Common Swift Apus apus annual breeding success may be related to weather conditions. Ornis Fenn. 83, 66–72.
Remisiewicz, M., Underhill, L.G., 2020. Climatic variation in Africa and Europe has combined effects on timing of spring migration in a long-distance migrant Willow Warbler Phylloscopus trochilus. PeerJ 8, e8770. .
Remisiewicz, M., Underhill, L.G., 2022. Large-scale climatic patterns have stronger carry-over effects than local temperatures on spring phenology of long-distance passerine migrants between Europe and Africa. Animals 12, 1732.
Romano, A., Garamszegi, L.Z., Rubolini, D., Ambrosini, R., 2023. Temporal shifts in avian phenology across the circannual cycle in a rapidly changing climate: a global meta-analysis. Ecol. Monogr. 93, e1552. .
Rubolini, D., Ambrosini, R., Caffi, M., Brichetti, P., Armiraglio, S., Saino, N., 2007. Longterm trends in first arrival and first egg laying dates of some migrant and resident bird species in northern Italy. Int. J. Biometeorol. 51, 553–563. https://link.springer.com/article/10.1007/s00484-007-0094-7.
Sæther, B.E., Sutherland, W.J., Engen, S., 2004. Climate influences on avian population dynamics. Adv. Ecol. Res. 35, 185–209. .
Salewski, V., Hochachka, W.M., Fiedler, W., 2013. Multiple weather factors affect apparent survival of European passerine birds. PLoS One 8, e59110. .
Shipley, J.R., Twining, C.W., Taff, C.C., Vitousek, M.N., Flack, A., Winkler, D.W., 2020. Birds advancing lay dates with warming springs face greater risk of chick mortality. Proc. Nat. Acad. Sci. USA 117, 25590–25594. .
Sicurella, B., Caffi, M., Caprioli, M., Rubolini, D., Saino, N., Ambrosini, R., 2015. Weather conditions, brood size and hatching order affect Common Swift Apus apus nestlings’ survival and growth. Bird Study 62, 64–77. .
Sparks, T.H., 1999. Phenology and the changing pattern of bird migration in Britain. Int. J. Biometeorol. 42, 134–138.
Stige, L.C., Stave, J., Chan, K.S., Ciannelli, L., Pettorelli, N., Glantz, M., et al., 2006. The effect of climate variation on agro-pastoral production in Africa. Proc. Nat. Acad. Sci. USA 103, 3049–3053. .
Taylor, C.M., Laughlin, A.J., Hall, R.J., 2016. The response of migratory populations to phenological change: a Migratory Flow Network modelling approach. J. Anim. Ecol. 85, 648–659. .
Thomson, D.L., Douglas-Home, H., Furness, R.W., Monaghan, P., 1996. Breeding success and survival in the Common Swift Apus apus: a long-term study on the effect of weather. J. Zool. 239, 29–38.
Tigges, U., 2008. Common swift worldwide. .
Trenberth, K.E., 1997. The definition of El Nino. Bull. Am. Meteorol. Soc. 78, 2771–2777.
Vahatalo, A.V., Rainio, K., Lehikoinen, A., Leikoinen, E., 2004. Spring arrival of birds depends on North Atlantic oscillation. J. Avian Biol. 35, 210–216. .
Verhulst, S., Nilsson, J.A., 2008. The timing of birds’ breeding seasons: a review of experiments that manipulated timing of breeding. Phil. Trans. R. Soc. B 363, 399–410. .
Visser, M.E., Both, C., 2005. Shifts in phenology due to global climate change: the need for a yardstick. Proc. R. Soc. B 272, 2561–2569. .
Visser, M.E., Adriaensen, F., van Balen, J.H., Blondel, J., Dhondt, A.A., van Dongen, S., et al., 2003. Variable responses to large-scale climate change in European Parus populations. Proc. R. Soc. B 270, 367–372.
Walker, W.H., Melendez-Fernandez, O.H., Nelson, R.J., Reiter, R.J., 2019. Global climate change and invariable photoperiods: a mismatch that jeopardizes animal fitness. Ecol. Evol. 9, 10044–10054. .
Walther, G.R., 2010. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B 365, 2019–2024. .
Weitnauer, E., 1980. Mein Vogel: aus dem Leben des Mauerseglers Apus apus. Basellandschaftlicher Vogelschutzverband, Oltingen.
Table
2.
Relationship between mean annual laying date (Julian day) in Roccamalatina and NAO or ENSO indices in the winter period, and local temperature or local rainfall during selected decades.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds