E Gaqie, Yuehong Cheng, Panyan Yang, Luhong Wang, Mei Xiao, Long Zhang, Caiquan Zhou, Pinjia Que, Bin Wang. 2025: Impacts of free-ranging yaks on habitat occupancy and population density of a high-mountain rare pheasant species. Avian Research, 16(1): 100238. DOI: 10.1016/j.avrs.2025.100238
Citation:
E Gaqie, Yuehong Cheng, Panyan Yang, Luhong Wang, Mei Xiao, Long Zhang, Caiquan Zhou, Pinjia Que, Bin Wang. 2025: Impacts of free-ranging yaks on habitat occupancy and population density of a high-mountain rare pheasant species. Avian Research, 16(1): 100238. DOI: 10.1016/j.avrs.2025.100238
E Gaqie, Yuehong Cheng, Panyan Yang, Luhong Wang, Mei Xiao, Long Zhang, Caiquan Zhou, Pinjia Que, Bin Wang. 2025: Impacts of free-ranging yaks on habitat occupancy and population density of a high-mountain rare pheasant species. Avian Research, 16(1): 100238. DOI: 10.1016/j.avrs.2025.100238
Citation:
E Gaqie, Yuehong Cheng, Panyan Yang, Luhong Wang, Mei Xiao, Long Zhang, Caiquan Zhou, Pinjia Que, Bin Wang. 2025: Impacts of free-ranging yaks on habitat occupancy and population density of a high-mountain rare pheasant species. Avian Research, 16(1): 100238. DOI: 10.1016/j.avrs.2025.100238
Chengdu Research Base of Giant Panda Breeding, Sichuan Key Laboratory of Conservation Biology for Endangered Wildlife, Chengdu, 610081, China. E-mail address: pinjia@panda.org.cn (P. Que)
Key Laboratory of Southwest China Wildlife Resources Conservation (Ministry of Education), China West Normal University, Nanchong, 637009, China. E-mail address: wangbin513@cwnu.edu.cn (B. Wang)
Free-ranging yak grazing is a regime specially adapted to high-elevation environments across the Pan-Tibetan Highlands, yet its impacts on alpine birds remain poorly understood. The Chinese Monal (Lophophorus lhuysii) is a rare pheasant species that serves as a representative and umbrella species for alpine meadow ecosystems in the mountains of Southwest China, and has long been regarded as threatened by free-ranging yaks. However, the actual impacts and specific mechanisms through which yak grazing influences Chinese Monal have not been empirically tested. Here, we conducted infrared camera monitoring in alpine meadows within the Wolong National Nature Reserve, Sichuan, China, from 2019 to 2022. We analyzed the effects of free-ranging yaks on habitat occupancy, activity pattern, and population density of the Chinese Monal using multispecies occupancy models, kernel density estimations, avoidance attraction ratios, and random encounter models. We found that interactions with yaks affect monal habitat occupancy in conjunction with other ecological factors. Specifically, the presence of yaks alters monals’ occupancy responses to variations in grass cover and elevation, causing monals to use habitats with lower grass cover and elevation more frequently. Additionally, the activity patterns of the Chinese Monal and yaks are significantly different, and the presence of yaks significantly prolongs the time until monals re-occupy the same habitats. As an outcome of the cumulative effects of spatial and temporal avoidance, the population density of the Chinese Monal negatively correlated with increasing grazing intensity. This study provides the first empirical evidence of the negative impacts of free-ranging yaks on the Chinese Monal and elucidates the underlying mechanisms, highlighting great risks to conservation of this vulnerable species. Our findings inform the optimization of grazing management that balances production with conservation. Strict control over yak numbers and grazing areas in critical habitats for Chinese Monal and other threatened species could be a feasible compromise to mitigate these pressures.
Livestock production, as one of the most significant and widespread forms of human land use, now occupies over 26% of the Earth’s terrestrial surface (FAO, 2017), and has had substantial and profound impacts on various natural ecosystems and biological communities (Filazzola et al., 2020; Maestre et al., 2022). With global population and food demand projected to increase significantly in the coming decades (Alexandratos and Bruinsma, 2012; OECD and FAO, 2020), the scope and intensity of these impacts are expected to expand and intensify, presenting unprecedented challenges for biodiversity conservation at both local and global scales (Phalan et al., 2011; Pozo et al., 2021). In this context, understanding the effects of livestock grazing on biodiversity and developing grazing regimes that balance food production with ecological conservation are crucial for mitigating the biodiversity crisis and achieving sustainable development goals (Wang and Tang, 2019; Filazzola et al., 2020; Pozo et al., 2021).
Birds are highly sensitive to environmental changes and disturbances, often serving as indicators of biodiversity and ecosystem health (Fraixedas et al., 2020). Over recent decades, the relationship between livestock grazing and sympatric bird species has become a prominent topic in conservation ecology and has been extensively studied across diverse regions, ecosystems, and taxa (Dettenmaier et al., 2017; Barzan et al., 2021; Wu et al., 2023). Previous studies suggested that livestock grazing can influence birds through multiple pathways, with impacts on habitat use as a primary mechanism (Fuller and Gough, 1999). However, the effects and mechanisms were highly variable; for instance, livestock may compete exploitatively with sympatric bird species when they share similar habitats or food resources, potentially excluding birds from optimal habitats (Schieltz and Rubenstein, 2016; Zarco et al., 2019). Livestock foraging and trampling can reduce understory cover and height, diminishing habitat occupancy of several bird species due to reduced habitat suitability or nesting site availability (Gonnet, 2001; Richmond et al., 2012; Poessel et al., 2020). In contrast, bird species preferring open habitats may benefit from lower and sparser understory resulting from grazing (Neilly and Schwarzkopf, 2019; Dardanelli et al., 2022; Nugent et al., 2022). As a consequence of these divergent responses, and possibly in combination with other factors, local populations of some bird species experienced significant declines under grazing pressure (Amar et al., 2011; Bellis and Muriel, 2015), while others coexist or even thrive with livestock grazing (García et al., 2008; Faria and Morales, 2020). Therefore, the impacts of livestock grazing on bird habitats and populations are highly dependent on environmental conditions, grazing regimes, and focal species (Schieltz and Rubenstein, 2016; Barzan et al., 2021), making it challenging to generalize findings across different systems. This underscores the need to uncover the interactions in specific livestock-wildlife systems, particularly when the goal is to conserve endangered and rare wildlife.
Free-ranging Yak (Bos grunniens, family Bovidae, order Artiodactyla) grazing is a unique regime specially adapted to high-elevation environments across the Pan-Tibetan Highlands, integral not only to the pastoral economy but also to the cultural and religious practices of local communities for several countries (Long et al., 2008; Wu et al., 2016). It has far-reaching impacts on fragile alpine meadow ecosystems, which are characterized by harsh environmental conditions, low productivity, sensitivity to climate change, and limited resilience and recovery (He et al., 2022). Previous researches have revealed multiple effects of yak grazing on plant diversity and productivity (e.g., Wang et al., 2018b; Mipam et al., 2019), as well as its varying relationships with wild ungulates (e.g., Mishra et al., 2004; Shrestha and Wegge, 2008) and carnivores (e.g., Shao et al., 2021). However, knowledge regarding its roles and implications for bird diversity conservation remains limited due to inadequate study effort compared to other grazing systems and taxa (Arthur et al., 2008), thereby hindering effective conservation of the rich diversity of endangered and endemic bird species in alpine meadow ecosystems (Dirnböck et al., 2011; Assandri et al., 2019).
The Chinese Monal (Lophophorus lhuysii, family Phasianidae, order Galliformes) is a rare and charismatic bird species endemic to China (Zheng, 2015). It is distributed exclusively in the high mountains of Southwest China, primarily inhabiting alpine meadows, shrubs, and screes at elevations of 3000–4900 m (Fig. 1A). It is listed as Vulnerable (VU) by the International Union for Conservation of Nature due to its limited and declining population (Birdlife International, 2022) and is included in Appendix A of Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES, 2024). The Chinese government has provided it legal protection as a Class Ⅰ key protected wild animal since 1989 (NFGA and MARA, 2021) and has included it in the ’14th Five-Year Plan’ for the rescue and conservation of rare and endangered wildlife (NFGA and NDRC, 2021). Additionally, it serves as a representative and umbrella species for alpine ecosystems (Wang et al., 2017; Shi et al., 2019). Across the mountains of Southwest China, the alpine meadows that the Chinese Monal depends on are also prime pastures for free-ranging yaks (Fig. 1B). During the summer and autumn, which coincide with the crucial breeding season of the Chinese Monal, large herds of yaks roam and graze in these meadows. As a result, the decline of the Chinese Monal has long been attributed to habitat degradation and disturbance caused by yak grazing (He et al., 1986; Lu et al., 1986; Ma, 1988; Long et al., 1998; Yu et al., 2017). However, no study has systematically tested this long-standing hypothesis.
Figure
1.
A typical alpine meadow habitat shared by the Chinese Monal (A, male) and free-ranging yak (B), photographed in the Wolong National Nature Reserve, Sichuan, China by an infrared camera.
In this study, we conducted extensive, long-term monitoring of the spatiotemporal dynamics of Chinese Monals and free-ranging yaks using infrared cameras in an alpine meadow ecosystem within the Wolong National Nature Reserve, Sichuan Province. Based on these monitoring data, we analyzed habitat use and interspecific interactions between the Chinese Monal and yak using multispecies occupancy models, assessed their temporal interactions using kernel density estimations and avoidance attraction ratios, and estimated their population densities across four subareas with varying grazing intensities using random encounter models. Our objective was to use the Chinese Monal as an indicator to reveal the impacts and underlying mechanisms of yak grazing on habitat and population dynamics of sympatric birds with similar ecological requirements, and to inform grazing management strategies for better conservation of alpine meadow biodiversity.
2.
Materials and methods
2.1
Study area
The Wolong National Nature Reserve (102°52′–103°24′ E, 30°45′–31°25′ N) is situated in southwestern Wenchuan County, Sichuan Province, China, covering an area of 2000 km2 and with elevations from 1150 m to 6250 m. The reserve lies on the eastern edge of the Qinghai-Tibet plateau climate zone, primarily characterized by a subtropical inland mountain climate. It receives an average annual sunshine duration of 926.7 h, an average annual temperature of 9.8 ℃, and an average annual precipitation of 1800 mm, with relative humidity exceeding 80%. The reserve is characterized by distinct vegetation zones that vary with elevation, including evergreen broad-leaved forests (<1600 m), evergreen-deciduous broad-leaved mixed forests (1600–2000 m), coniferous-broadleaf mixed forests (2000–2600 m), coniferous forests (2600–3800 m), alpine shrubs and meadows (3800–4400 m), and sparse alpine vegetation on screes (4400–5000 m). The alpine meadows are often important pastures for grazing yaks (Hull et al., 2014).
2.2
Infrared camera monitoring
We conducted two sessions of infrared camera monitoring in the study area. First, to examine the effects of yak grazing on the habitat use and activity pattern of the Chinese Monal, we conducted monitoring from April 2019 to September 2020. We used ArcGIS 10.6 (ESRI, Redlands, CA, USA) to divide the study area into 500 m × 500 m grids. Considering vegetation, site accessibility, and the monal’s elevation range, we hierarchically sampled 84 grids at elevations from 3420 to 4962 m. In each of the 84 grids, we deployed one infrared camera (L710, Yianws, Shenzhen, China), ensuring a minimum distance of over 300 m between adjacent camera sites (Fig. 2A) (Li et al., 2012). Cameras were mounted 0.5–1.0 m above ground on trunks or rocks, with grasses and shrubs cleared to reduce false triggers. Each camera was set to capture three photos and a 10 s video per trigger, with a 30 s interval and medium sensor sensitivity. Cameras operated continuously, with batteries and memory cards replaced approximately every three months.
Figure
2.
Infrared camera site for monitoring the Chinese Monal and free-ranging yaks during 2019–2020 (A) and 2022 (B) in alpine meadows in the Wolong National Nature Reserve. There is a wire fence along the ridge between Maanqiao and Muxiangpo for dividing pastures, preventing the yaks to cross.
Subsequently, to assess the impact of yak grazing on monal population, we selected four subareas with similar vegetation and habitat characteristics but varying grazing intensities: Cuomo (no grazing), Maanqiao (low grazing intensity), Muxiangpo (moderate grazing intensity), and Balangshan (high grazing intensity). We randomly sampled 44 grids (500 m × 500 m) across these subareas (11 in Cuomo, 10 in Maanqiao, 11 in Muxiangpo, and 12 in Balangshan) and deployed one camera in each grid, ensuring a minimum distance of over 300 m between adjacent camera sites (Fig. 2B). We conducted the second monitoring session from June to September 2022, with the same camera setup as those of the first monitoring session in 2019–2020.
2.3
Occupancy modeling
We applied a multispecies occupancy model to investigate occupancy patterns and interspecific relationships between the Chinese Monal and yaks. This modeling approach predicts marginal occupancy probabilities of individual species independently, as well as conditional occupancy probabilities of a target species in the presence or absence of another species, enabling rigorous testing of interspecific relationships (asymmetric competition, neutral coexistence, or mutual attraction) (Rota et al., 2016). Additionally, it quantifies the relative contributions of interspecific interactions and environmental covariates on occupancy probability, thereby identifying the key factors influencing species occupancy dynamics. Using data from the first monitoring session (2019–2020), we generated detection history matrices for the Chinese Monal and yaks, focusing only on the monal’s breeding season (April–September) when both species share alpine meadow habitats. We divided the monitoring session into 20 15-day sampling periods, recording any detection of the target species (monals or yaks) at a given site in a given period as "1" and non-detection as "0". Missing data due to camera malfunction or loss were recorded as "NA". To address the issue of imperfect detection (MacKenzie et al., 2002), we included monitoring effort (measured by the number of operational days for each camera per sampling period) as a candidate detection covariate, as detection rates for a target species are expected to correlate positively with monitoring effort (Andrade-Ponce et al., 2022).
We selected 11 ecological factors potentially influencing habitat use of the Chinese Monal and yak as candidate occupancy covariates. Habitat features, including shrub cover, average shrub height, grass cover, average grass height, and rock cover, were estimated using 10 m × 10 m quadrats centered on each camera. Trees were rare throughout the alpine meadows, and was negligible in our quadrats; thus, we did not include related features. We downloaded Digital Elevation Model (DEM) data with a 90 m resolution from the Geospatial Data Cloud (http://www.gscloud.cn) to generate raster layers of elevation, aspect, and slope in ArcGIS 10.6. Aspect was transformed from a circular to a linear variable ranging from −1 to 1 using a cosine function, to indicate north-facing (1, shady) and south-facing (−1, sunny) slopes (Wang et al., 2023). We obtained a river vector layer from the National Catalogue Service for Geographic Information (https://www.webmap.cn) and used it to generate a raster layer of Euclidean distance to the nearest river in ArcGIS 10.6. Elevation, aspect, slope, and distance to the nearest river were then extracted to each camera site based on coordinates.
Based on the literatures (Lu et al., 1986; Wang et al., 2008, 2018a; Bu et al., 2019), we identified potential predators of the Chinese Monal detected in the infrared camera monitoring, classifying them into nest predators (primarily targeting eggs and chicks) and individual predators (targeting adult birds) (Appendix Table S1). To measure predation risk for monals, we calculated a Relative Abundance Index (RAI) for each predator group at each camera site:
RAIi=NiTrapdayi×100
(1)
where Ni represents the total independent captures of nest or individual predators at site i, and Trapdayi is the total camera-trap days at site i. Multiple photos or videos of the same species at the same site within 30 min were considered as one independent capture (O’Brien et al., 2003).
To facilitate numerical optimization and avoid parameter estimation failures, we normalized all occupancy covariates to a 0–1 range using min-max normalization (Gu et al., 2022). We assessed multicollinearity by calculating Pearson correlation coefficients (r) for each covariate pair and retained only one covariate where |r| ≥ 0.6 (Tan et al., 2017). Consequently, we retained aspect, slope, elevation, distance to the nearest river, shrub cover, grass cover, average grass height, nest predator RAI, and individual predator RAI as occupancy covariates, while rock cover and average shrub height were excluded due to high correlation (Appendix Fig. S1).
To streamline the multispecies occupancy model, we employed a two-step modeling approach to optimize the combination of detection and occupancy covariates (Andrade-Ponce et al., 2022). First, we constructed single-species occupancy models for the Chinese Monal, including its detection history matrix, nine occupancy covariates, and one detection covariate. Akaike Information Criterion values corrected for small samples (AICc) and model weights were calculated to evaluate models with different covariate combinations. Covariates within optimal models (ΔAICc ≤2) were considered factors influencing habitat occupancy (Burnham and Anderson, 2002) and were included in subsequent multispecies occupancy modeling. This procedure was repeated for yaks to select their influential covariates. Next, these selected occupancy and detection covariates were incorporated into a set of multispecies occupancy models. To explore interactions between the Chinese Monal and yaks, we tested three hypotheses about interspecific interactions in the multispecies occupancy models: Hypothesis 1 assumes independent effects of occupancy covariates on monal and yak occupancy, with no interspecific interaction; Hypothesis 2 posits that occupancy probabilities are influenced by both covariates and interspecific interactions, where interactions between species are constant; Hypothesis 3 suggests that occupancy probabilities depend on both covariates and interspecific interactions, with species responses to occupancy covariates varying with the presence or absence of the other species. For Hypothesis 3, we created a model for each combination of occupancy covariates interacting with the presence or absence of the other species. We calculated AICc value for each model and inferred occupancy patterns and interspecific interactions based on the hypothesis and covariates of the model with the lowest AICc value.
Additionally, we evaluated the relative importance of interspecific interactions and ecological factors in influencing species occupancy. The odds ratio associated with the presence or absence of the other species (ORsp) when covariate h is at a baseline value x was calculated as follows:
ORsp=odds(z1=1∣z2=1,h=x)odds(z1=1∣z2=0,h=x)
(2)
Similarly, the odds ratio associated with a change in covariate h of Δx units (ORh) was calculated as:
Based on the monitoring data from 2019 to 2020, we used kernel density estimation to analyze the daily activity patterns of the Chinese Monal and yak. First, the "overlap" package (Ridout and Linkie, 2009) in R was used to generate the activity rhythms of both species and calculate the coefficient of overlap (Δ). The range of this coefficient is from 0 (no overlap) to 1 (complete overlap). Given that the sample sizes for both species were greater than 50, the Δ4 method was employed to compute the overlap coefficient Δ, and the 95% confidence intervals were calculated through 1000 bootstrap iterations (Meredith et al., 2024). Subsequently, to determine the interspecific difference in activity pattern, a Wald test with 1000 iterations was performed to compare probabilistic distributions of the activity rhythms of the two species, using "activity" package (Rowcliffe, 2023). A P < 0.05 was considered to indicate a significant difference.
To further explore the temporal response of the Chinese Monal to the presence of yak, we calculated the avoidance attraction ratios (AAR) for each camera trap location based on the detection history matrix of the two species:
AARi=T2iT1i
(4)
where T1i is the average length of time between successive monal detections without any intervening yak detections at site i, and T2i represents the same interval but with yak detections in between (Parsons et al., 2016). An AAR value equal to 1 indicates a neutral response of the Chinese Monal to yak presence, while values greater or less than 1 suggest temporal avoidance or attraction, respectively. We then applied a one-sample t-test to determine if the observed AAR differed significantly from 1.
2.5
Population density estimation
To examine the effects of grazing intensity on Chinese Monal population, we used Random Encounter Models (REM) (Rowcliffe et al., 2008) to estimate population densities of the Chinese Monal and yaks in each of the four subareas, based on monitoring data from the second session in 2022:
D=yt×πvr(2+θ)×g
(5)
where D is population density of the target species in a subarea, y is the number of independent captures of the target species, t is the total number of camera-trap days, v represents the average daily movement distance of the target species, g represents the average group size of the target species, and r and θ are the radius and angle of the camera detection zone, respectively.
REM estimates animal density based on species-specific characteristics and camera trap detection parameters, without requiring individual identification or accounting for spatial autocorrelation between camera sites and animal movement. An essential parameter required in this model is the target species’ average daily movement distance, ideally calculated using satellite tracking data (Pettigrew et al., 2021). For the Chinese Monal, we tracked two adult individuals (one male and one female) in the Tangjiahe National Nature Reserve, Sichuan, over 31 days during a breeding season using backpack satellite trackers (HQZN, Global Messenger, Hunan, China), accumulating 462 valid activity points (P. Que, unpubl. data). The average daily movement distance was calculated as 1.334 km/day. For yak movement distance, we referenced the findings of Gu et al. (2023), who tracked yaks on the Tibetan plateau using GPS collars, estimating an average daily movement of 9.073 km/day. Based on the cameras’ product specifications and our measurements in the study area, we set the detection radius to 10 m (i.e., r = 0.01 km) and the detection angle to 40° (i.e., θ = 0.698 radians). The Chinese Monal and yaks are not solitary and were often photographed in groups; thus, independent captures represent groups rather than individuals. In such cases, the density estimate D reflects group density, which we converted to individual density by multiplying it by an unbiased estimate of the average group size (g) for each target species (Rowcliffe et al., 2008). We calculated the average group size for each species in each subarea by summing the number of individuals in all independent captures throughout the monitoring session, then dividing by the total number of independent captures (Rademaker et al., 2016). Since these estimates of daily movement distance are based on daytime activity, we only used independent captures of both species between 06:00 and 20:00 to estimate population density. Finally, we calculated a Pearson correlation coefficient between estimated population densities to examine the effect of grazing intensity on the Chinese Monal population. An α-priori of P < 0.05 was considered significant.
3.
Results
3.1
Species habitat occupancy and interspecific interaction
During the first monitoring session (2019–2020), the total monitoring effort amounted to 17,903 camera-trap days over the breeding seasons. We detected 59 animal species with 7509 independent captures. The Chinese Monal was recorded at 56 sites with 599 independent captures, while free-ranging yaks were recorded at 31 sites with 570 independent captures. In total, we detected 18 potential predator species of the Chinese Monal, comprising nine potential nest predators and nine potential individual predators (Appendix Table S1). Human activities were rare throughout the study area, and only recorded at five sites with 11 independent captures.
Single-species occupancy models indicated five optimal models (ΔAICc ≤2) for the Chinese Monal and nine for yaks (Table 1). Five and six ecological factors were identified as influencing habitat occupancy for the Chinese Monal and yaks, respectively: slope, distance to the nearest river, shrub cover, and grass cover were shared factors, aspect and elevation influenced only yaks, while RAI of individual predators affected only the Chinese Monal (Table 1). In addition, monitoring effort was found to influence the detection probability of yaks only.
Table
1.
Optimal single species occupancy models for the Chinese Monal and free-ranging yak evaluated based on Akaike Information Criterion corrected for small sample sizes (AICc).
Model structure
K
Log-likelihood
AICc
ΔAICc
Weight
Chinese Monal
P (.); Ψ(DTR + SLO)
4
−585.115
1178.736
0
0.308
P (.); Ψ(DTR + GRC + SLO)
5
−584.325
1179.418
0.682
0.219
P (.); Ψ(DTR + PAI + SLO)
5
−584.440
1179.649
0.912
0.195
P (.); Ψ(DTR + GRC + PAI + SLO)
6
−583.530
1180.151
1.415
0.152
P (.); Ψ(DTR + SHC + SLO)
5
−584.875
1180.520
1.783
0.126
Yak
P (.); Ψ(GRC + SHC + SLO)
5
−341.356
693.480
0
0.199
P (.); Ψ(DTR + GRC + SHC + SLO)
6
−340.633
694.357
0.877
0.128
P (.); Ψ(DTR + ELE + GRC + SHC + SLO)
7
−339.453
694.380
0.899
0.127
P (.); Ψ(ELE + GRC + SHC + SLO)
6
−340.798
694.686
1.206
0.109
P (.); Ψ(GRC + SHC)
4
−343.187
694.881
1.400
0.099
P (.); Ψ(ASP + GRC + SHC + SLO)
6
−340.934
694.958
1.478
0.095
P (.); Ψ(SHC + SLO)
4
−343.324
695.153
1.673
0.086
P (.); Ψ(DTR + GRC + SHC)
5
−342.222
695.212
1.732
0.084
P (DAY); Ψ(GRC + SHC + SLO)
6
−341.170
695.430
1.950
0.075
P(): detection covariates; Ψ(): occupancy covariates; K: Number of parameters; DAY: number of days in operation for each camera site in each sampling period; ASP: aspect; SLO: slope; ELE: elevation; DTR: distance to the nearest river; SHC: shrub cover; GRC: grass cover; PAI: relative abundance index of predators potentially preying Chinese Monal individuals.
Using these covariates, we constructed 129 multispecies occupancy models under three hypotheses on interspecific interaction (one for Hypothesis 1, one for Hypothesis 2, and 127 for Hypothesis 3; Appendix Table S2). The top model with the lowest AICc fell under Hypothesis 3, where occupancy probabilities of the Chinese Monal and yaks were influenced by their respective occupancy covariates and interspecific interaction, with their responses to grass cover and elevation interacting with this interspecific interaction (Appendix Table S2). The marginal occupancy probability of the Chinese Monal was positively corelated with slope, distance to the nearest river, grass cover, and RAI of individual predators, while negatively correlated with shrub cover (Fig. 3). For yaks, the marginal occupancy probability was negatively correlated with slope and distance to the nearest river, and positively correlated with shrub cover, grass cover, aspect, and elevation (Fig. 3).
Figure
3.
Marginal occupancy probability of the Chinese Monal and free-ranging yaks along gradients of important ecological factors. ASP: aspect; SLO: slope; ELE: elevation; DTR: distance to the nearest river; SHC: shrub cover; GRC: grass cover; PAI: relative abundance index of predators potentially preying Chinese Monal individuals. Lines show posterior means and shaded regions are 95% confidence intervals.
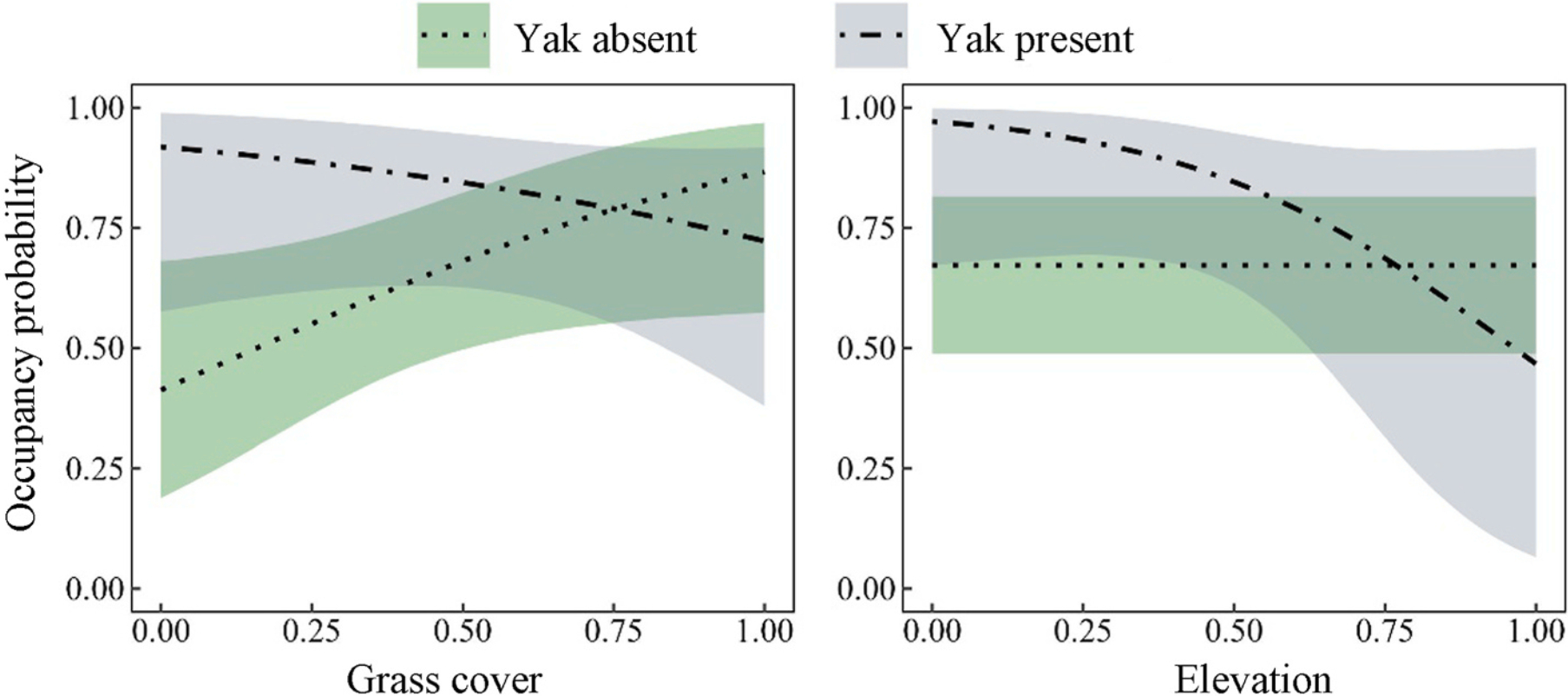
The conditional occupancy predicted by the top multispecies occupancy model suggested an interspecific interaction coupled with grass cover and elevation. Specifically, the probability of Chinese Monal occupancy was positively correlated with grass cover when yak was absent, but the presence of yaks reversed this into a negative correlation, with a relatively lower co-occurrence probability in habitats with higher grass cover (Fig. 4). Similarly, Chinese Monal occupancy probability was invariant with elevation in the absence of yaks, consistent with the pattern observed in marginal occupancy where elevation was not an influencing factor (Fig. 3), but a negative correlation emerged when yaks were present (Fig. 4). Additionally, the absolute log odds ratio of the interaction between the Chinese Monal and yaks (0.997) was greater than other occupancy covariates, suggesting it as the most important factor affecting Chinese Monal occupancy (Appendix Table S3).
Figure
4.
Occupancy probability of the Chinese Monal conditional on the presence and absence of free-ranging yaks along gradients of grass cover and elevation. Lines show posterior means and shaded regions are 95% confidence intervals.
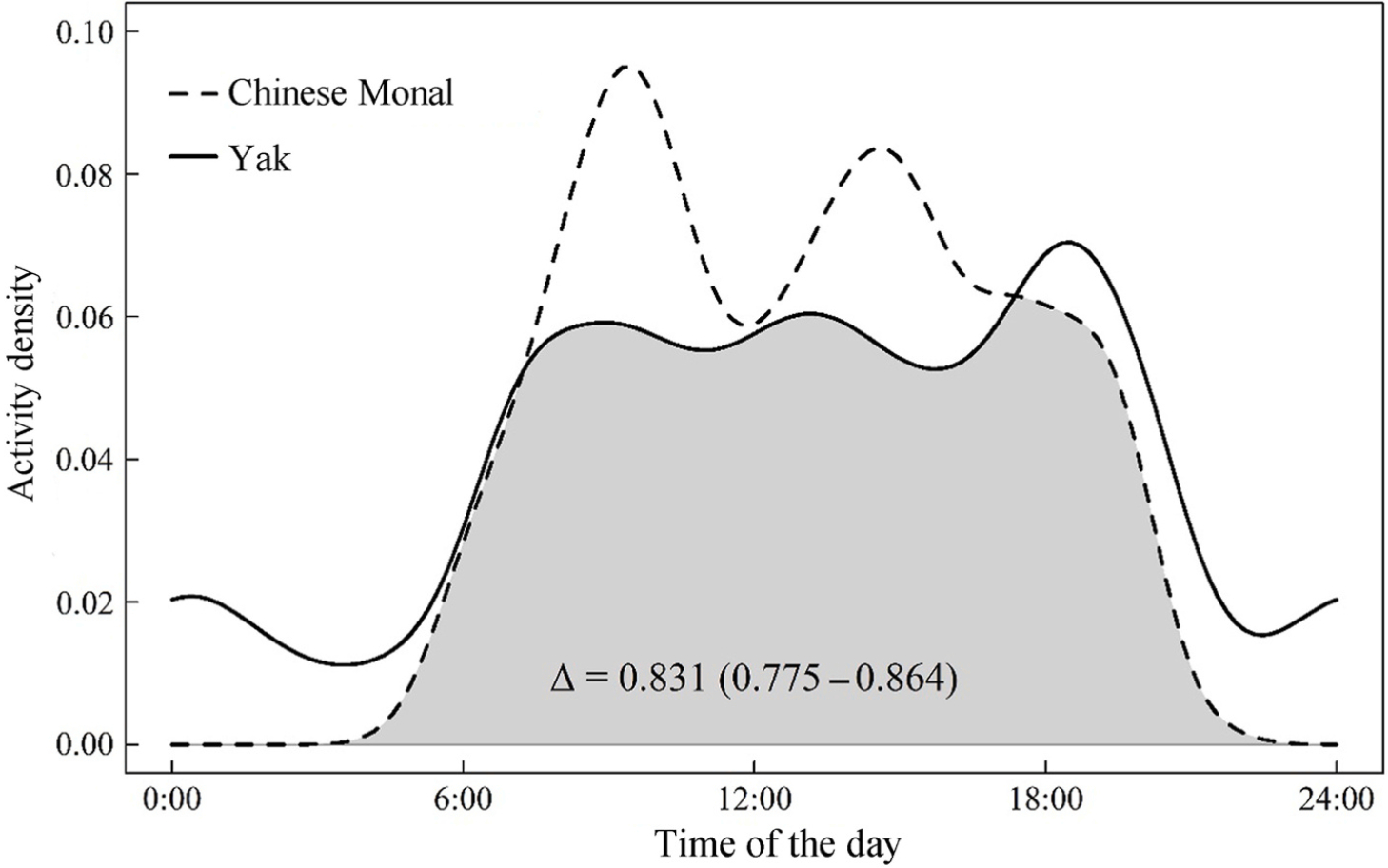
The analysis of daily activity patterns indicated that the Chinese Monal is a typical diurnal species, with a bimodal activity pattern. The activity peaks occurred around 09:30 in the morning and 14:30 in the afternoon, respectively. In contrast, although the yaks mainly active during daytime, with peak occurring around 18:00 in the dusk, they performed several behaviors at night as well. The temporal overlap coefficient between the Chinese Monal and yak was estimated as 0.831 (95% CI: 0.775–0.864) (Fig. 5), and the Wald test indicated a significant difference in their activity patterns (P < 0.001).
Figure
5.
Daily activity patterns and temporal activity overlap coefficient of the Chinese Monal and yak.
Additionally, we found evidence of behavioral avoidance by the Chinese Monal in response to free-ranging yaks. The mean AAR averaged across the 14 camera sites where both T1 and T2 were calculable was 4.178 ± 0.930 and significantly greater than 1 (one-sample t-test, df = 13, t = 3.415, P = 0.002). This result indicates that when yaks were present, the intervals between successive monal detections increased, suggesting that yaks’ presence prolonged the time until monals reappeared in the same habitats.
3.3
Population density
In the second monitoring session (2022), the total monitoring effort amounted 4404 camera-trap days across the four subareas with varying grazing intensities. The total number of daytime independent captures of the Chinese Monal and free-ranging yaks was 132 and 357, respectively. Estimates based on the REMs revealed differences in population densities for both the Chinese Monal and yaks across the subareas. Yak density estimates aligned with the expected grazing intensities: Balangshan had the highest yak density, followed by Muxiangpo and Maanqiao, with no yaks recorded in Cuomo (Table 2). The estimated population density of the Chinese Monal was negatively, but not significantly, correlated with yak density (r = −0.895, P = 0.105). In the ungrazed subarea of Cuomo, Chinese Monal density was over 2.5 times higher than in the heavily grazed subarea of Balangshan (Table 2).
Table
2.
Population density estimations of the Chinese Monal and free-ranging yaks in the four subareas with varying grazing intensity in Wolong National Nature Reserve.
Habitat occupancy of the Chinese Monal and free-ranging yaks in alpine meadows is influenced by various ecological factors, many of which affect the two species in contrasting ways. Our occupancy model indicates that the Chinese Monal prefers habitats with lower shrub cover, aligning with previous findings (Yang et al., 2022). Field observations suggested that shrubs can provide the Chinese Monal concealment from predators (Zhang, 1995) and roosting sites (He et al., 1986). However, as a large, ground-foraging bird, the Chinese Monal may find dense shrubs obstructive to movement and foraging, which may explain its avoidance of shrubby habitats. Conversely, yaks strongly prefer shrubby habitats, as shrubs like Salicaceae and Rosaceae species contribute substantially to their summer and autumn diets (Teng et al., 2013, 2014). Our results show that the Chinese Monal’s habitat occupancy is positively correlated with distance from rivers, which matches previous ecological niche modeling studies (Zhong et al., 2021). Chinese Monals typically inhabit high elevations, relying on smaller water sources, like rain puddles and seasonal streams, rather than perennial rivers. However, yaks, as large livestock, likely have a greater need for water, leading them to prefer riverine areas (Zhao et al., 2012).
We found that yak occupancy is highly correlated with topographic factors, with a strong preference for shaded slopes at higher elevations with gentle slopes. Large body size of yaks hindered their roaming on steep terrain, making gentle areas more suitable for their foraging (Liu et al., 2014). Additionally, shaded slopes are typically more humid, promoting denser plant growth, which provide ample food resources for yaks (Xiao et al., 2019). In contrast, the Chinese Monal shows a preference for steeper habitats, aligning with previous findings (Zhang, 1995; Zhou et al., 2022). This is likely due to their anti-predation strategies and breeding behaviors. Limited in flight, they rely on steep terrain to glide away from predators (Li et al., 2023). During courtship, males also use elevated sites for calling and display behaviors, such as gliding from rocks or treetops, making steep slopes ideal for these displays (He et al., 1986). Interestingly, we found a positive correlation between Chinese Monal occupancy and predation pressure. While typically prey species avoid high-predation-risk areas, Chinese Monals may be unable to actively avoid these habitats due to the spatial restrictions of home range during breeding period (e.g., courtship, incubation, and chick-rearing), leaving them exposed. Predators, in turn, tend to select habitats where prey is more abundant, reinforcing this observed correlation (Karanth et al., 2011; Dou et al., 2019).
In addition to these ecological factors that have contrasting effects, grass cover influences the Chinese Monal and yaks similarly under average conditions, with both showing a general preference for higher grass cover. This finding is unsurprising, as both species rely mainly on herbaceous plants and prefer grassy habitats as a means to access richer food resources. However, interspecific interactions alter these dynamics. In the absence of yaks, conditional occupancy of the Chinese Monal remains positively correlated with grass cover, consistent with its marginal habitat occupancy. But the presence of yaks reverses this relationship to a negative correlation. Dietary overlap between the Chinese Monal and yaks during the breeding season—particularly herbaceous species in the families Brassicaceae, Poaceae, and Ranunculaceae—suggests potential competition for food resources (Teng et al., 2013, 2014; Wang et al., 2022). Thereby, although both species prefer grassy habitats with richer food resource, yaks likely exclude the Chinese Monal from optimal habitats through exploitative competition (Wang et al., 2015). Yak occupancy’s relationship with grass cover remains little affected by the presence of the Chinese Monal (Appendix Fig. S2), further underscoring the asymmetry of this competition. Similarly, elevation interacts with interspecific interactions. The Chinese Monal’s marginal occupancy remains unaffected by elevation within the study range (3420–4962 m). However, yak presence forces the Chinese Monal to lower elevations more frequently. These findings provide strong evidence of spatial avoidance of yaks by the Chinese Monal, with yak influence outweighing other ecological factors and proving decisive in habitat selection of the Chinese Monal. Additionally, regarding temporal interaction, both analyses of daily activity patterns and avoidance attraction ratios suggest a significantly temporal avoidance of the Chinese Monal in response to yaks.
Our results largely differ from previous studies in other pheasant-livestock systems. For instance, livestock presence and abundance were found to positively correlate with habitat occupancy of the Blood Pheasant (Ithaginis cruentus) (Fan et al., 2020) and the Green Peafowl (Pavo muticus) (Gu et al., 2022). Neither species exhibited temporal avoidance to livestock (Fan et al., 2020; Gu et al., 2022), while the Reeves’s Pheasant (Syrmaticus reevesii) altered behavioral patterns in response to grazing (Hua et al., 2022). These divergent effects are likely due to different ecosystems (alpine meadows for our study vs. mountainous forests for the three cases), differing livestock species (yak vs. cattle, horses, and goat), and distinct pheasant species, emphasizing the uniqueness of free-ranging yaks in alpine meadow ecosystems. In addition, the multispecies occupancy model used in our study can incorporate dynamics of both the Chinese Monal and yaks to analyze marginal and conditional occupancy, having advantages over single-species occupancy models in disentangling the effects of interspecific interactions from decoupled factors (Rota et al., 2016). Moreover, using the consistent monitoring and REMs, we found a negative correlation between Chinese Monal and yak population densities. This supports earlier speculation that human activities, such as grazing and medicinal herbs collection, resulted in regional differences in Chinese Monal population density (Yu et al., 2017). Therefore, the negative relationship between Chinese Monal population density and grazing intensity we highlighted across different subareas likely reflects the cumulative effects of both spatial and temporal avoidance mechanisms, and highlights a potential long-term risk to the species in heavily grazed areas.
This study represents the first attempt to systematically test the effects of free-ranging yak grazing on the Chinese Monal. While we acknowledge limitations in sample size, monitoring intensity, and the scope of impact dimensions assessed (e.g., predation risk, food resource, and breeding ecology), our findings nevertheless reveal significant grazing-induced threats to this vulnerable species and elucidate the underlying ecological mechanisms. These results establish a critical foundation for future research and offer immediate insights for conservation managements. The continuing increase in yak grazing intensity in high-elevation areas (Wu et al., 2016; Chai et al., 2019) will inevitably exacerbate the threats to the Chinese Monal. However, given the importance of yak grazing to the local pastoral economy, a complete grazing ban is impractical (Zhang et al., 2017). Therefore, based on our findings, strict control over yak numbers and grazing areas in critical habitats for the Chinese Monal and other species facing similar threats could serve as a feasible compromise to mitigate the pressure posed by yak grazing on alpine biodiversity conservation. Moreover, a multi-dimensional and multi-scale integrated research framework is essential for the conservation of the Chinese Monal and the development of sustainable grazing practices. Future studies should prioritize three key areas: (1) the direct effects of yak grazing on breeding success and food resource availability of the Chinese Monal; (2) indirect impacts mediated through altered monals’ interspecific interactions with competitors and predators; and (3) finer-scale influences on monal microbiology and parasitology.
CRediT authorship contribution statement
Gaqie E: Writing – original draft, Visualization, Investigation, Formal analysis. Yuehong Cheng: Resources, Investigation, Data curation. Panyan Yang: Data curation. Luhong Wang: Investigation, Data curation. Mei Xiao: Investigation, Data curation. Long Zhang: Data curation. Caiquan Zhou: Project administration. Pinjia Que: Writing – review & editing, Investigation, Funding acquisition. Bin Wang: Writing – review & editing, Funding acquisition, Conceptualization.
Ethics statement
The experiments reported here comply with the current laws of China. The capture and tracking of the Chinese Monal was carried out with specific permit of National Forestry and Grassland Administration (forestry permit [2023] 175).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank Arielle Parsons and Yu Xu for their help in model interpretation, as well as the staff from Wolong National Nature Reserve and Tangjiahe National Nature Reserve for their assistance in fieldwork.
Alexandratos, N., Bruinsma, J., 2012. World agriculture towards 2030/2050: the 2012 revision. In: UN Food and Agricultural Organization. FAO, Rome.
Amar, A., Davies, J., Meek, E., Williams, J., Knight, A., Redpath, S., 2011. Long-term impact of changes in sheep Ovis aries densities on the breeding output of the hen harrier Circus cyaneus. J. Appl. Ecol. 48, 220–227.
Andrade-Ponce, G.P., Mandujano, S., Dáttilo, W., Farías-González, V., Jiménez, J., Velásquez-C, K., et al., 2022. A framework to interpret co-occurrence patterns from camera trap data: the case of the gray fox, the bobcat, and the eastern cottontail rabbit in a tropical dry habitat. J. Zool. 318, 91–103.
Arthur, A.D., Pech, R.P., Davey, C., Jiebu, Zhang, Y., Lin, H., 2008. Livestock grazing, plateau pikas and the conservation of avian biodiversity on the Tibetan plateau. Biol. Conserv. 141, 1972–1981.
Assandri, G., Bogliani, G., Pedrini, P., Brambilla, M., 2019. Species-specific responses to habitat and livestock management call for carefully targeted conservation strategies for declining meadow birds. J. Nat. Conserv. 52, 125757.
Bartoń, K., 2024. MuMIn: multi-model inference. R package version 1.48.4. . (Accessed 11 July 2024).
Barzan, F.R., Bellis, L.M., Dardanelli, S., 2021. Livestock grazing constrains bird abundance and species richness: a global meta-analysis. Basic Appl. Ecol. 56, 289–298.
Bellis, L.M., Muriel, N., 2015. Response of the endemic Long-tailed Meadowlark (Sturnella loyca obscura) to grazing exclusion in herbivore-dependent upland grasslands of Argentina. Emu 115, 176–184.
Birdlife International, 2022. Lophophorus lhuysii. The IUCN Red List of Threatened Species 2022: e.T22679192A219003994 (Accessed 29 June 2023).
Bu, H., Shen, X., Li, S., 2019. Predation patterns on artificial nests of ground nesting pheasants in the montane forest, Southwest China. Acta Ornithol. 54, 35–43, 9.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach. Springer-Verlag, New York.
Chai, J., Yu, X., Xu, C., Xiao, H., Zhang, J., Yang, H., et al., 2019. Effects of yak and Tibetan sheep trampling on soil properties in the northeastern Qinghai-Tibetan plateau. Appl. Soil Ecol. 144, 147–154.
CITES, 2024. Convention on international trade in endangered species of wild Fauna and Flora (CITES) appendices Ⅰ, Ⅱ and Ⅲ. .
Dettenmaier, S.J., Messmer, T.A., Hovick, T.J., Dahlgren, D.K., 2017. Effects of livestock grazing on rangeland biodiversity: a meta-analysis of grouse populations. Ecol. Evol. 7, 7620–7627.
Dirnböck, T., Essl, F., Rabitsch, W., 2011. Disproportional risk for habitat loss of high‐altitude endemic species under climate change. Glob. Change Biol. 17, 990–996.
Dou, H., Yang, H., Smith, J.L.D., Feng, L., Wang, T., Ge, J., 2019. Prey selection of Amur tigers in relation to the spatiotemporal overlap with prey across the Sino-Russian border. Wildl. Biol. 2019 wlb.00508.
Fan, F., Bu, H., Mcshea, W.J., Shen, X., Li, B.V., Li, S., 2020. Seasonal habitat use and activity patterns of blood pheasant Ithaginis cruentus in the presence of free-ranging livestock. Global Ecol. Conserv. 23, e01155.
FAO, 2017. The State of Food and Agriculture: leveraging food systems for inclusive rural transformation. .
Faria, N., Morales, M.B., 2020. Livestock species and grazing rotational patterns modulate grassland bird assemblages in Mediterranean drylands. Agric. Ecosyst. Environ. 295, 106893.
Filazzola, A., Brown, C., Dettlaff, M.A., Batbaatar, A., Grenke, J., Bao, T., et al., 2020. The effects of livestock grazing on biodiversity are multi-trophic: a meta-analysis. Ecol. Lett. 23, 1298–1309.
Fiske, I.J., Chandler, R.B., 2011. unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Software 43, 1–23.
Fraixedas, S., Lindén, A., Piha, M., Cabeza, M., Gregory, R., Lehikoinen, A., 2020. A state-of-the-art review on birds as indicators of biodiversity: advances, challenges, and future directions. Ecol. Indic. 118, 106728.
Fuller, R.J., Gough, S.J., 1999. Changes in sheep numbers in Britain: implications for bird populations. Biol. Conserv. 91, 73–89.
García, C., Renison, D., Cingolani, A.M., Fernández-Juricic, E., 2008. Avifaunal changes as a consequence of large-scale livestock exclusion in the mountains of Central Argentina. J. Appl. Ecol. 45, 351–360.
Gonnet, J.M., 2001. Influence of cattle grazing on population density and species richness of granivorous birds (Emberizidae) in the arid plain of the Monte, Argentina. J. Arid Environ. 48, 569–579.
Gu, B., Weng, Y., Diao, Y., Zhao, Q., Zhang, Z., Tian, S., et al., 2022. Is livestock grazing compatible with green peafowl (Pavo muticus) conservation? Potential chance of peafowl-human coexistence. Biol. Conserv. 275, 109772.
Gu, C., Liu, L., Zhang, Y., Wei, B., Cui, B., Gong, D., 2023. Understanding the spatial heterogeneity of grazing pressure in the Three-River-Source Region on the Tibetan plateau. J. Geogr. Sci. 33, 1660–1680.
He, F.Q., Lu, T.X., Lu, C.L., Cui, X.Z., 1986. Study on the breeding ecology of the Chinese Monal. Acta Ecol. Sin. 6, 186–192.
He, Y.L., Wang, J.S., Tian, D.S., Quan, Q., Jiang, L., Ma, F.F., et al., 2022. Long-term drought aggravates instability of alpine grassland productivity to extreme climatic event. Ecology 103, e3792.
Hua, J., Lu, S., Song, K., Wang, J., Wang, J., Xu, J., 2022. Effects of livestock grazing on spatio-temporal patterns and behaviour of Reeves’s pheasant Syrmaticus reevesii. Animals 12, 2968.
Hull, V., Zhang, J., Zhou, S., Huang, J., Viña, A., Liu, W., et al., 2014. Impact of livestock on giant pandas and their habitat. J. Nat. Conserv. 22, 256–264.
Karanth, K.U., Gopalaswamy, A.M., Kumar, N.S., Vaidyanathan, S., Nichols, J.D., Mackenzie, D.I., 2011. Monitoring carnivore populations at the landscape scale: occupancy modelling of tigers from sign surveys. J. Appl. Ecol. 48, 1048–1056.
Kellner, K.F., Smith, A.D., Royle, J.A., Kery, M., Belant, J.L., Chandler, R.B., 2023. The unmarked R package: twelve years of advances in occurrence and abundance modelling in ecology. Methods Ecol. Evol. 14, 1408–1415.
Li, J., Peng, B., Li, D., He, F., Zheng, B., Lin, Y., et al., 2023. Report on the breeding habits of Lophophorus lhuysii in the xiaozhaizigou national nature reserve, sichuan. Sichuan J. Zool. 42, 601–608.
Li, S., Mcshea, W.J., Wang, D., Lu, Z., Gu, X., 2012. Gauging the impact of management expertise on the distribution of large mammals across protected areas. Divers. Distrib. 18, 1166–1176.
Liu, Z., Sun, Z., Li, L., Zhao, C., Li, Y., Wang, Y., 2014. Habitat selection of feral yak in different time periods in Helan Mountains, China. J. Northeast For. Univ. 42, 73–77.
Long, R.J., Ding, L.M., Shang, Z.H., Guo, X.H., 2008. The yak grazing system on the Qinghai-Tibetan plateau and its status. Rangel. J. 30, 241–246.
Long, T.L., Shao, K.Q., Guo, G., Cheng, C.Y., Zou, X.Y., Landel, H., et al., 1998. Field tracking and ecological observation of the Chinese Monal in winter. Sichuan J. Zool. 17, 104–105.
Lu, T.C., Liu, R.S., He, F.Q., Lu, C.L., 1986. Ecological studies on Chinese monal. Acta Zool. Sin. 32, 273–279.
Ma, G.Y., 1988. Observation on the Chinese monal in gansu Province. Sichuan J. Zool. 7, 41–42.
MacKenzie, D.I., Nichols, J.D., Lachman, G.B., Droege, S., Royle, J.A., Langtimm, C.A., 2002. Estimating site occupancy rates when detection probabilities are less than one. Ecology 83, 2248–2255.
Maestre, F.T., Le Bagousse-Pinguet, Y., Delgado-Baquerizo, M., Eldridge, D.J., Saiz, H., Berdugo, M., et al., 2022. Grazing and ecosystem service delivery in global drylands. Science 378, 915–920.
Meredith, M., Ridout, M., Campbell, L.A., 2024. overlap: estimates of coefficient of overlapping for animal activity patterns. R package version 0.3.9. . (Accessed 26 November 2024).
Mipam, T. -D., Zhong, L. -L., Liu, J. -Q., Miehe, G., Tian, L. -M., 2019. Productive overcompensation of alpine meadows in response to yak grazing in the eastern Qinghai-Tibet plateau. Front. Plant Sci. 10, 925.
Mishra, C., Van Wieren, S.E., Ketner, P., Heitkönig, I.M.A., Prins, H.H.T., 2004. Competition between domestic livestock and wild bharal Pseudois nayaur in the Indian Trans-Himalaya. J. Appl. Ecol. 41, 344–354.
Neilly, H., Schwarzkopf, L., 2019. The impact of cattle grazing regimes on tropical savanna bird assemblages. Austral Ecol. 44, 187–198.
NFGA (National Forestry and Grassland Administration), MARA (Ministry of Agriculture and Rural Affairs of the People’s Republic of China), 2021. The List of National Key Protected Wild Animal Species. Beijing, China.
NFGA (National Forestry and Grassland Administration), NDRC (National Development and Reform Commission), 2021. The Outline of the National 14th Five-Year Plan on the Conservation and Development of Forestry and Grassland. Beijing, China.
Nugent, D.T., Baker-Gabb, D.J., Leonard, S.W.J., Morgan, J.W., 2022. Livestock grazing to maintain habitat of a critically endangered grassland bird: is grazer species important? Ecol. Appl. 32, e2587.
O’Brien, T.G., Kinnaird, M.F., Wibisono, H.T., 2003. Crouching tigers, hidden prey: sumatran tiger and prey populations in a tropical forest landscape. Anim. Conserv. 6, 131–139.
OECD, FAO, 2020. OECD-FAO Agricultural Outlook 2020–2029 (Summary). OECD Publishing.
Parsons, A.W., Bland, C., Forrester, T., Baker-Whatton, M.C., Schuttler, S.G., Mcshea, W.J., et al., 2016. The ecological impact of humans and dogs on wildlife in protected areas in eastern North America. Biol. Conserv. 203, 75–88.
Parsons, A.W., Rota, C.T., Forrester, T., Baker‐Whatton, M.C., Mcshea, W.J., Schuttler, S.G., et al., 2019. Urbanization focuses carnivore activity in remaining natural habitats, increasing species interactions. J. Appl. Ecol. 56, 1894–1904.
Pettigrew, P., Sigouin, D., St-Laurent, M. -H., 2021. Testing the precision and sensitivity of density estimates obtained with a camera-trap method revealed limitations and opportunities. Ecol. Evol. 11, 7879–7889.
Phalan, B., Onial, M., Balmford, A., Green, R.E., 2011. Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science 333, 1289–1291.
Poessel, S.A., Hagar, J.C., Haggerty, P.K., Katzner, T.E., 2020. Removal of cattle grazing correlates with increases in vegetation productivity and in abundance of imperiled breeding birds. Biol. Conserv. 241, 108378.
Pozo, R.A., Cusack, J.J., Acebes, P., Malo, J.E., Traba, J., Iranzo, E.C., et al., 2021. Reconciling livestock production and wild herbivore conservation: challenges and opportunities. Trends Ecol. Evol. 36, 750–761.
R Core Team, 2024. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. .
Rademaker, M., Meijaard, E., Semiadi, G., Blokland, S., Neilson, E.W., Rode-Margono, E.J., 2016. First ecological study of the Bawean warty pig (Sus blouchi), one of the rarest pigs on Earth. PLoS One 11, e0151732.
Richmond, O.M.W., Tecklin, J., Beissinger, S.R., 2012. Impact of cattle grazing on the occupancy of a cryptic, threatened rail. Ecol. Appl. 22, 1655–1664.
Ridout, M.S., Linkie, M., 2009. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 14, 322–337.
Rota, C.T., Ferreira, M.A.R., Kays, R.W., Forrester, T.D., Kalies, E.L., Mcshea, W.J., et al., 2016. A multispecies occupancy model for two or more interacting species. Methods Ecol. Evol. 7, 1164–1173.
Rowcliffe, J.M., Field, J., Turvey, S.T., Carbone, C., 2008. Estimating animal density using camera traps without the need for individual recognition. J. Appl. Ecol. 45, 1228–1236.
Rowcliffe, M., 2023. activity: animal activity statistics. R package version 1.3.4. . (Accessed 26 November 2024).
Schieltz, J.M., Rubenstein, D.I., 2016. Evidence based review: positive versus negative effects of livestock grazing on wildlife. What do we really know? Environ. Res. Lett. 11, 113003.
Shao, X.N., Lu, Q., Xiong, M.Y., Bu, H.L., Shi, X.Y., Wang, D.J., et al., 2021. Prey partitioning and livestock consumption in the world’s richest large carnivore assemblage. Curr. Biol. 31, 4887–4897.
Shi, X.W., Gong, C., Zhang, L., Hu, J., Ouyang, Z.Y., Xiao, Y., 2019. Which species should we focus on? Umbrella species assessment in Southwest China. Biology 8, 42.
Shrestha, R., Wegge, P., 2008. Habitat relationships between wild and domestic ungulates in Nepalese trans-Himalaya. J. Arid Environ. 72, 914–925.
Tan, C.K.W., Rocha, D.G., Clements, G.R., Brenes-Mora, E., Hedges, L., et al., 2017. Habitat use and predicted range for the mainland clouded leopard Neofelis nebulosa in Peninsular Malaysia. Biol. Conserv. 206, 65–74.
Teng, L.W., Li, Y.X., Liu, Z.S., Dong, J.P., Yao, Z.C., Zhang, J.T., 2013. Summer diet composition of feral yak in Helan Mountains. J. Econ. Anim. 17, 192–196.
Teng, L.W., Wang, Y., Li, L.Y., Yao, Z.C., Lu, P.F., Li, Y.X., et al., 2014. Autumn diet composition of feral yak (Bos grunnieus) in Helan Mountains. J. Anhui Agr. Sci. 42, 6258–6260.
Wang, B., Li, Y., Zhang, G., Yang, J., Deng, C., Hu, H., et al., 2022. Seasonal variations in the plant diet of the Chinese monal revealed by fecal DNA metabarcoding analysis. Avian Res. 13, 100034.
Wang, B., Xu, Y., Ran, J.H., 2017. Predicting suitable habitat of the Chinese monal (Lophophorus lhuysii) using ecological niche modeling in the Qionglai Mountains, China. PeerJ 5, e3477.
Wang, B., Xu, Y., Zhang, B., Wu, Y.J., He, X.C., Ran, J.H., et al., 2018a. Overlap and selection of dust-bathing sites among three sympatric montane galliform species. Auk 135, 1076–1086.
Wang, B., Zhong, X., Xu, Y., Cheng, Y., Ran, J., Zhang, J., et al., 2023. Optimizing the Giant Panda National Park’s zoning designations as an example for extending conservation from flagship species to regional biodiversity. Biol. Conserv. 281, 109996.
Wang, C., Tang, Y., 2019. A global meta-analyses of the response of multi-taxa diversity to grazing intensity in grasslands. Environ. Res. Lett. 14, 114003.
Wang, F., Mcshea, W.J., Wang, D., Li, S., 2015. Shared resources between giant panda and sympatric wild and domestic mammals. Biol. Conserv. 186, 319–325.
Wang, J., Zhang, C., Yang, H., Mou, C., Mo, L., Luo, P., 2018b. Plant community ecological strategy assembly response to yak grazing in an alpine meadow on the eastern Tibetan plateau. Land Degrad. Dev. 29, 2920–2931.
Wang, W., Ma, J., Yu, H., Hu, L., 2008. Food habits of asiatic black bears in the xiao xing’anling mountains. Acta Theriol. Sin. 28, 7–13.
Wickham, H., 2016. ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York, USA.
Wu, N., Yi, S., Srijana, J., Neha, B., 2016. Yak on the Move: Transboundary Challenges and Opportunities for Yak Raising in a Changing Hindu Kush Himalayan Region. International Centre for Integrated Mountain Development, Nepal.
Wu, Y., Du, Y., Liu, X., Wan, X., Yin, B., Hao, Y., et al., 2023. Grassland biodiversity response to livestock grazing, productivity, and climate varies across biome components and diversity measurements. Sci. Total Environ. 878, 162994.
Xiao, X., Gericaidan, Hou, F.J., 2019. Effects of grazing and topography on α diversity and physical properties of the soil of alpine meadow communities in the Qinghai-Tibet plateau. Pratacult. Sci. 36, 3041–3051.
Yang, H.L., Cheng, Y.H., Zhou, T.X., Feng, X., Hu, Q., Zhang, G.Q., et al., 2022. Multi-scale habitat selection of Chinese monal (Lophophorus lhuysii) in Wolong national nature reserve, sichuan. Biodivers. Science 30, 21535.
Yu, X., Chen, J., Wang, B., Yan, Y., Ran, J., He, F., et al., 2017. Population density estimation and habitat suitability assessment of Lophophorus lhuysii during breeding season in Xiaozhaizigou National Nature Reserve, Sichuan Province. Sichuan J. Zool. 36, 361–367.
Zarco, A., Cueto, V.R., Sagario, M.C., Marone, L., 2019. Effects of livestock grazing on flocks of seed-eating birds in the central Monte desert, Argentina. Can. J. Zool. 97, 606–611.
Zhang, J., Hull, V., Ouyang, Z., Li, R., Connor, T., Yang, H., et al., 2017. Divergent responses of sympatric species to livestock encroachment at fine spatiotemporal scales. Biol. Conserv. 209, 119–129.
Zhang, T., 1995. Distribution and ecology of Chinese monals in baishuijiang nature reserve, gansu. Chin. J. Zool. 30, 25–28.
Zhao, C.N., Su, Y., Liu, Z.S., Yao, Z.C., Zhang, M.M., Li, Z.G., 2012. Habitat selection of feral yak in winter and spring in the Helan Mountains, China. Acta Ecol. Sin. 32, 1762–1772.
Zheng, G.M., 2015. Pheasants in China. Higher Education Press, Beijing.
Zhong, X., Yang, N., Zhang, L., Cheng, Y.H., Feng, X., Hu, Q., et al., 2021. Population survey and habitat quality assessment of Lophophorus lhuysii in the Wolong national nature reserve. Sichuan J. Zool. 40, 509–516.
Zhou, T.X., Yang, H.L., Zhang, G.Q., Yang, J., Feng, X., Hu, Q., et al., 2022. Temporal and spatial niche differentiation among three alpine Galliformes with sympatric distribution in the Wolong National Nature Reserve, Sichuan Province. Biodivers. Sci. 30, 22026.
Table
1.
Optimal single species occupancy models for the Chinese Monal and free-ranging yak evaluated based on Akaike Information Criterion corrected for small sample sizes (AICc).
Model structure
K
Log-likelihood
AICc
ΔAICc
Weight
Chinese Monal
P (.); Ψ(DTR + SLO)
4
−585.115
1178.736
0
0.308
P (.); Ψ(DTR + GRC + SLO)
5
−584.325
1179.418
0.682
0.219
P (.); Ψ(DTR + PAI + SLO)
5
−584.440
1179.649
0.912
0.195
P (.); Ψ(DTR + GRC + PAI + SLO)
6
−583.530
1180.151
1.415
0.152
P (.); Ψ(DTR + SHC + SLO)
5
−584.875
1180.520
1.783
0.126
Yak
P (.); Ψ(GRC + SHC + SLO)
5
−341.356
693.480
0
0.199
P (.); Ψ(DTR + GRC + SHC + SLO)
6
−340.633
694.357
0.877
0.128
P (.); Ψ(DTR + ELE + GRC + SHC + SLO)
7
−339.453
694.380
0.899
0.127
P (.); Ψ(ELE + GRC + SHC + SLO)
6
−340.798
694.686
1.206
0.109
P (.); Ψ(GRC + SHC)
4
−343.187
694.881
1.400
0.099
P (.); Ψ(ASP + GRC + SHC + SLO)
6
−340.934
694.958
1.478
0.095
P (.); Ψ(SHC + SLO)
4
−343.324
695.153
1.673
0.086
P (.); Ψ(DTR + GRC + SHC)
5
−342.222
695.212
1.732
0.084
P (DAY); Ψ(GRC + SHC + SLO)
6
−341.170
695.430
1.950
0.075
P(): detection covariates; Ψ(): occupancy covariates; K: Number of parameters; DAY: number of days in operation for each camera site in each sampling period; ASP: aspect; SLO: slope; ELE: elevation; DTR: distance to the nearest river; SHC: shrub cover; GRC: grass cover; PAI: relative abundance index of predators potentially preying Chinese Monal individuals.
Table
2.
Population density estimations of the Chinese Monal and free-ranging yaks in the four subareas with varying grazing intensity in Wolong National Nature Reserve.


 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds