Evaluation of antimicrobial resistance and virulence characteristics in Escherichia coli and Proteus mirabilis isolates from dead Crested Ibis embryos at the artificial breeding base
Bacterial infections of avian embryos can lead to an increase in embryo mortality, and the proliferation of antimicrobial-resistant bacteria aggravates the situation. A low hatching rate also poses a challenge to the population of artificially bred Crested Ibises (Nipponia nippon). This study aims to determine the potential association between bacterial infection and the death of Crested Ibis embryos, and whether there is convergence between antimicrobial resistance and virulence in strain. In this study, 13 Escherichia coli and 12 Proteus mirabilis isolates were recovered from dead Crested Ibis embryos. The pathogenicity examination confirmed the pathogenicity of all isolates, and multiple virulence genes detected by PCR-sequencing demonstrated the presence of irp2 and iucD (100%), fimC and iss (92.31%) in E. coli, and ucaA (58.33%) in P. mirabilis. Antimicrobial susceptibility test demonstrated that isolates were mainly resistant to amoxicillin (E. coli: 76.92%, P. mirabilis: 91.67%), cefazolin (E. coli: 76.92%, P. mirabilis: 91.67%), oxytetracycline (E. coli: 92.31%, P. mirabilis: 75.00%) and sulfamethoxazole-trimethoprim (E. coli: 53.85%, P. mirabilis: 33.33%), and more than 30% of isolates showed multidrug-resistance (MDR). Further analyses detected extended-spectrum β-lactamase (ESBL) genes, of which blaTEM-1 (E. coli: 100%, P. mirabilis: 100%) had the highest frequency, followed by the blaCTX-M-55 (E. coli: 92.31%, P. mirabilis: 50%), blaCTX-M-14 (E. coli: 76.92%, P. mirabilis: 33.33%), blaCTX-M-65 (E. coli: 15.38%, P. mirabilis: 16.67%), and all isolates were negative for blaSHV and blaOXA. Pearson’s correlation analysis showed a positive correlation between the presence of β-lactam resistance and ESBL genes, while mainly negative correlations were observed between the presence of ESBL genes and virulence genes. Furthermore, the conjugation experiment and PFGE revealed that the isolates were primarily polyclonal, and there was horizontal transfer of resistance or virulence genes by plasmids. Based on the results, E. coli and P. mirabilis were responsible for embryonic mortality of the ibises in this study. The co-presence and co-transfer of ESBL genes and virulence genes can pose a potential threat to the health of the Crested Ibis, and measures such as prudent use of antimicrobials, and constant surveillance of resistance and pathogenicity, must be implemented at the Crested Ibis breeding base.
Bacterial infections are generally considered the primary cause of embryonic death in poultry, with common pathogens including Escherichia coli, Bacillus cereus, Staphylococci spp., and Proteus spp. These bacteria can migrate into the egg yolk, ultimately leading to embryonic death (Karunarathna et al., 2022). E. coli and P. mirabilis are common Gram-negative bacteria found in the gastrointestinal tract of humans and animals, as well as in the environment (soil, water and sewage, etc.). Pathogenic strains of these bacteria can cause intestinal or extraintestinal diseases (Armbruster et al., 2018; Poirel et al., 2018). It has been reported that E. coli is a major cause of yolk sac infection, omphalitis, and early mortality in young chicks (Xi et al., 2001), while P. mirabilis has been implicated in reproductive failure in avian species (Rezaee et al., 2021). Currently, in clinical practice, the prevention and treatment of bacterial diseases primarily rely on the use of antimicrobials. However, due to genomic modifications driven by extensive use of antimicrobials and horizontal gene transfer through mobile elements (plasmids, integrons, IS, ICEs, etc.), multidrug-resistant strains have increasingly emerged, including those producing ESBLs, AmpC, carbapenemases and other antimicrobial resistances, making infections difficult to treat (Poirel et al., 2018; Girlich et al., 2020).
The Crested Ibis (Nipponia nippon) is one of the most endangered birds, classified as threatened by the International Union for Conservation of Nature (IUCN) Red List (Lu et al., 2022). Once widely distributed across East Asia, its population experienced a dramatic decline in the 20th century due to overhunting, habitat loss, and the use of agrochemicals. In 1981, the last seven wild Crested Ibises were discovered in Yangxian County, Shaanxi Province, China. In response, the Chinese government implemented a series of conservation measures to protect the species, leading to a gradual recovery in both population size and distribution (Fu et al., 2019; Yang et al., 2020; Lu et al., 2022). Despite these efforts, the Crested Ibis continues to face significant survival challenges, with high embryonic mortality being one of the most pressing concerns (Fu et al., 2019). A survey conducted at the Crested Ibis breeding base in Sichuan Province of China reported that the majority of embryonic deaths occurred during the late developmental stages, with the deceased embryos exhibiting symptoms such as small body size, poor yolk absorption, subcutaneous edema on the head, and curled toes (Yang et al., 2020). Moreover, primary health challenges during the breeding and brooding phase were characterized by intestinal, respiratory and ocular infections, which responded favorably to antimicrobial therapy. However, pathogens of death and diseases during breeding and brooding period were not analyzed in this report (Yang et al., 2020). A study reported by Xi et al. (2007) revealed that infectious diseases emerged rapidly at the Shaanxi Crested Ibis Breeding Center since 2000. During two months in 2000, six nestlings died with similar clinical symptoms such as anorexia, diarrhea, lameness and sudden death. The deaths, analyzed by autopsy and bacterial examination, were caused by E. coli strains. However, there have been no reports on whether E. coli or other bacterial infections are responsible for embryonic death in Crested Ibises. This study aimed to explore the pathogenicity of E. coli and P. mirabilis isolates from dead Crested Ibis embryos. In addition, antimicrobial resistance patterns and ESBL and virulence-associated genes were analyzed to determine whether convergence between virulence and resistance exists, which may pose a potential threat to the artificial breeding of the Crested Ibis.
2.
Materials and methods
2.1
Sampling, necropsy and bacteriologic examination
Between April 2020 and April 2021, a total of 23 samples of dead Crested Ibis embryos were collected from different parents at the Mount Emei Biological Resources and Experiment Station Base, Sichuan, China (Fig. 1). All samples were transported to the laboratory in a cooler box, and necropsies were performed immediately under sterile conditions. Liver samples were taken for bacteriological examination.
Figure
1.
Filed photos collected from the Crested Ibis breeding base. (A) Crested Ibis living in the base; (B) Hatching eggs of Crested Ibis; (C) The dead embryo of Crested Ibis in late incubation.
Liver samples were inoculated on sheep blood agar and MacConkey agar and incubated at 37 ℃ for 24 h. Gram staining, standard biochemical tests, and 16S rDNA PCR sequencing were performed (El-Tayeb et al., 2017).
2.2
Assessment of mouse pathogenicity
This study was approved by the Institutional Animal Care and Use Committee of Sichuan Agricultural University (permit No. 20200336). Five KM mice were inoculated by intraperitoneal injection (i.p.) with each strain in 0.2 mL culture (1×108 CFU per mouse). Control mice were administered 0.2 mL of sterile saline i.p. The challenged mice were observed three times daily for deaths. Strains causing at least one death within five days were designated pathogenic (Takeuchi et al., 2003).
2.3
Antimicrobial susceptibility test
Antimicrobial susceptibility test was performed using the broth microdilution method. The antimicrobials tested included amoxicillin (AMX), cefazolin (CZO), ceftriaxone (CRO), streptomycin (STR), gentamicin (GM), colistin sulfate (COL), oxytetracycline (OTC), nalidixic acid (NA), ciprofloxacin (CIP), and sulfamethoxazole-trimethoprim (SXT). Interpretative breakpoints were based on CLSI (Clinical and Laboratory Standards Institute) criteria (CLSI, 2021). MDR was defined as resistance to three or more antimicrobial categories.
2.4
Detection of extended-spectrum β-lactamase (ESBLs) genes and virulence genes by PCR-sequencing
Plasmid DNA was extracted using a plasmid mini kit (OMEGA) following kit instructions. Plasmid DNA was used as a template of PCR for the detection of the main ESBL resistance genes, including blaTEM, blaSHV, blaCTX-M-1 group, blaCTX-M-2 group, blaCTX-M-8/25 group, blaCTX-M-9 group and blaOXA (Table 1). The total DNA of strains was extracted using the boiling method and used as a template of PCR for virulence genes, including E. coli virulence genes (papC, tsh, fimC, hlyE, iss, iucD, and irp2) (Meng et al., 2014; Paixão et al., 2016; Hussain et al., 2022), and P. mirabilis virulence genes (rsbA, ureC, pmfA, atfA, zapA, ucaA, mrpA, and ptA), following the methods described in previous studies (Sanches et al., 2019; Yu et al., 2020; Algammal et al., 2021).
Table
1.
Sequences of PCR primers, target genes and amplicon sizes.
All of PCR products were sequenced by TSINGKE Biological Technology Co., Ltd (Chengdu, China), and the results were analyzed using the BLAST program and the available database on the National Center for Biotechnology Information (NCBI) website.
2.5
Plasmid conjugation experiment
Plasmid conjugation experiments were performed with the recipient strain E. coli J53 AZr to assess the transferability of resistance and virulence genes from 13 E. coli and 12 P. mirabilis donors. The transconjugants were selected in TSA agar plates containing azide (200 μg/mL) and amoxicillin (64 μg/mL) for 18–24 h. The PCR-sequencing and antimicrobial susceptibility test were used to examine the acquired resistance/virulence genes and resistance patterns of these transconjugants, respectively (Tran et al., 2021).
2.6
Pulsed field gel electrophoresis (PFGE) testing
Molecular typing of 13 E. coli and 12 P. mirabilis isolates was performed using PFGE, following the standardized protocol for PFGE from PulseNet USA and a previous report (Gautom, 1997). Briefly, agarose-embedded genomic DNA samples were digested with Xba I restriction enzyme (Takara, Japan), and the restricted fragments were separated in 1% agarose using the Chef Mapper pulse-field gel electrophoresis system. The Salmonella serotype Braenderup H9812 was used as a reference and molecular marker.
The PFGE patterns were analyzed using Bionumerics 7.6 software and the Dice coefficient. A dendrogram was generated using the unweighted pair-group method with arithmetic means (UPGMA). A study reported that strains with more than 80% similarity were considered to belong to the same clonal lineage (cluster) (Aghamohammad et al., 2022).
2.7
Statistical analysis
The Pearson’s correlation (r) between β-lactam resistance phenotype, ESBLs resistance genes, and virulence genes among isolates was analyzed. The presence of individual antibiotic resistance, ESBLs resistance gene or virulence gene received a score of 1, whereas susceptibility to antibiotics and the absence of ESBLs or virulence gene received a score of 0. Binary data (0/1) were imported into R software (4.2.2), the “cor” and “corrplot” function determined correlation and provided visualization, and the “cor. mtest” function was used for significance of correlation (P < 0.05). Strong correlation was considered if r ≥ 0.6, moderate correlation if r was between 0.4 and 0.6, and weak if r < 0.4 (Elbediwi et al., 2021).
3.
Results
3.1
Necropsy, bacteriology and pathogenicity examination
Anatomical examination of the dead embryos revealed that they were small, had curled toes, showed yolk sac inflammation, yolk malabsorption, and had bleeding spots on their body surfaces in some cases. In the bacteriological examination, 13 E. coli and 12 P. mirabilis isolates were successfully recovered, with both E. coli and P. mirabilis being isolated simultaneously from five samples of embryos.
In the assessment of mouse pathogenicity, mice showed symptoms such as depression, anorexia, eye inflammation, twitching, and labored respiration within 6 h post-infection. Most deaths occurred within 12 h post-infection, and all mice challenged died within 72 h. Necropsies of infected mice demonstrated slight hemorrhages in the lungs, swelling of some organs (liver, spleen, and kidney), and thin fluid- and gas-filled small intestines. No deaths or pathological changes were observed in the control group.
3.2
Antimicrobial susceptibility test
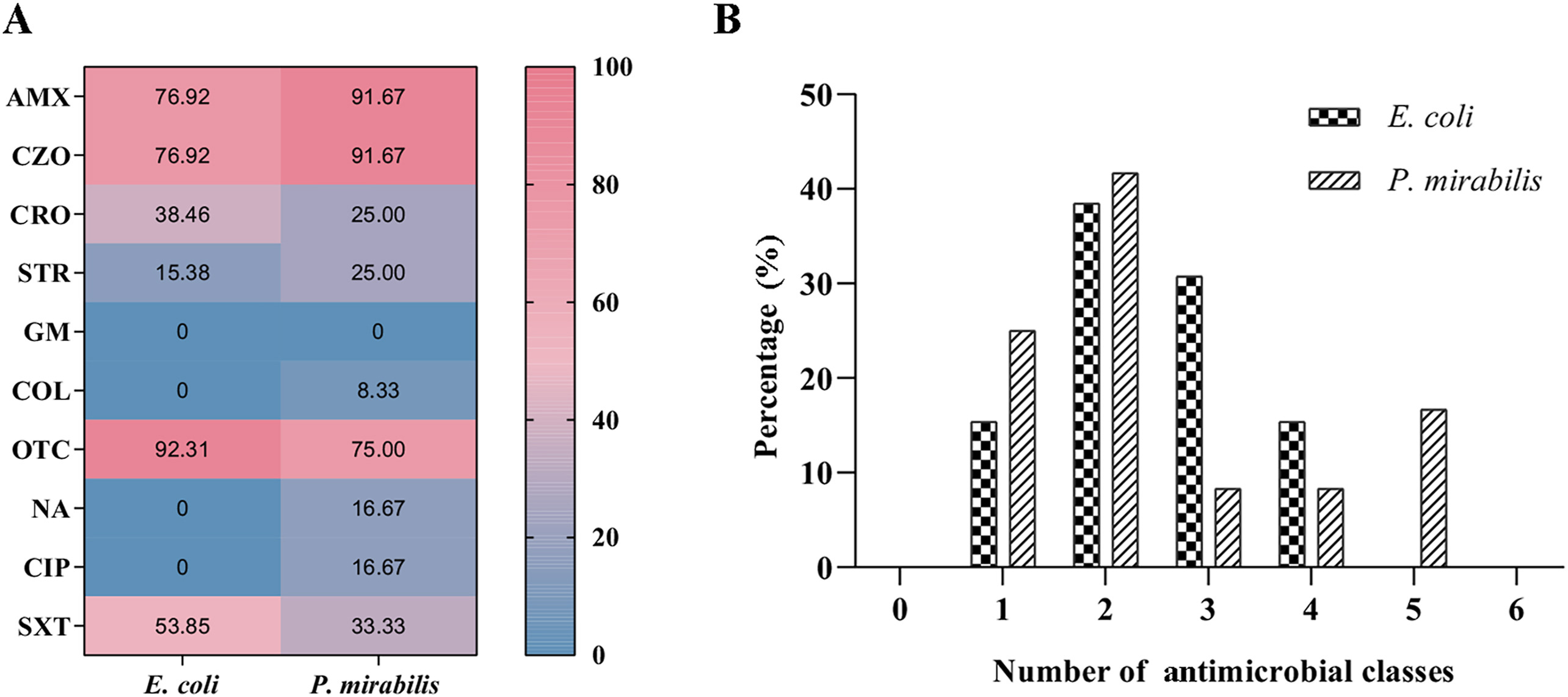
Antimicrobial susceptibility test showed that all isolates were resistant to at least one antimicrobial. Except for gentamicin, resistance to the other antimicrobials tested varied in frequency. There was particularly high resistance to amoxicillin (E. coli: 76.92%, P. mirabilis: 91.67%), cefazolin (E. coli: 76.92%, P. mirabilis: 91.67%), oxytetracycline (E. coli: 92.31%, P. mirabilis: 75.00%) and SXT (E. coli: 53.85%, P. mirabilis: 33.33%) (Fig. 2A), and 46.15% of E. coli and 33.33% of P. mirabilis were MDR (Fig. 2B).
Figure
2.
Antimicrobial resistance patterns among E. coli and P. mirabilis isolates. (A) Frequency of resistance among E. coli and P. mirabilis isolates; (B) The distributions of MDR among E. coli and P. mirabilis isolates.
3.3
Detection of ESBLs resistance genes and virulence genes
The results of PCR-sequencing of ESBLs genes revealed that all isolates carried blaTEM-1 (E. coli: 100%, P. mirabilis: 100%), followed by the blaCTX-M-55 (E. coli: 92.31%, P. mirabilis: 50%), blaCTX-M-14 (E. coli: 76.92%, P. mirabilis: 33.33%) and blaCTX-M-65 (E. coli: 15.38%, P. mirabilis: 16.67%). No blaSHV and blaOXA genes were detected. Additionally, 100% of E. coli and 75% of P. mirabilis isolates simultaneously carried multidrug ESBLs genes (Table 2).
Table
2.
The distributions of ESBLs resistance/virulence genes among E. coli and P. mirabilis isolates.
Among the 13 E. coli isolates, four virulence genes were detected, with the highest prevalence of irp2 and iucD (100%), followed by fimC and iss (92.31%). All isolates carried three or four virulence genes simultaneously. However, only ucaA (58.33%) was detected in the 12 P. mirabilis isolates. Notably, the co-presence of ESBL and virulence genes was detected in more than half of the strains (100% of E. coli, 58.33% of P. mirabilis) (Table 2).
3.4
Association of β-lactam resistance phenotype, ESBLs gene, and virulence gene among isolates
Correlation analysis of antimicrobial resistance phenotype, antimicrobial resistance gene, and virulence gene among the examined isolates was performed. All observed correlation results were statistically significant (P < 0.05) as indicated. For E. coli, there was a weak or moderate positive correlation between β-lactam resistance and ESBLs genes, such as cefazolin and blaCTX-M-14 (r = 0.13), ceftriaxone and blaCTX-M-55 (r = 0.23), ceftriaxone and blaCTX-M-14 (r = 0.43), and amoxicillin and blaCTX-M-14 (r = 0.57). Except for a moderate positive correlation between blaCTX-M-14 and iss (r = 0.53), there were negative correlations between blaCTX-M-14 and fimC (r = -0.16), blaCTX-M-65 and iss (r = -0.69).
For P. mirabilis, there was a moderate or strong positive correlation between β-lactam resistance and resistance genes, such as amoxicillin and blaCTX-M-55 (r = 0.3), cefazolin and blaCTX-M-55 (r = 0.3), ceftriaxone and blaCTX-M-55 (r = 0.58), ceftriaxone and blaCTX-M-65 (r = 0.77). There were negative correlations between each ESBLs gene and ucaA (r value ranged from -0.53 to -0.12), blaCTX-M-55 and ucaA (r = -0.51), blaCTX-M-65 and ucaA (r = -0.53).
3.5
Transfer ability of ESBLs and virulence genes
Conjugation experiments successfully produced ten and eleven transconjugants from the 13 E. coli and 12 P. mirabilis isolates, respectively. The PCR-sequencing results of related genes in the transconjugants revealed that 100% of blaTEM-1, 100% of blaCTX-M-55, 77.78% of blaCTX-M-14 and 70% of iss in E. coli donors were transferred to E. coli J53 AZr. For P. mirabilis, 100% of blaTEM-1, 66.67% of blaCTX-M-55, 66.67% of blaCTX-M-14, 100% of blaCTX-M-65 and 85.71% of ucaA in donors were also transferred.
The antimicrobial susceptibility test showed that the resistance of these transconjugants to antimicrobials tested significantly improved (Table 3). Furthermore, no significant difference was observed in the MDR rate between the donor bacteria (E. coli: 50.00%, P. mirabilis: 36.36%) and their respective transconjugants (E. coli transconjugants: 30.00%, P. mirabilis transconjugants: 18.18%) (P > 0.05) (Fig. 3).
Table
3.
The antimicrobial resistance patterns and virulence profiles of donor bacteria and their transconjugants.
Figure
3.
Comparison of the MDR rate between the donor bacteria and their respective transconjugants. (A) The E. coli donors (n = 10) and their transconjugants (P = 0.83); (B) The P. mirabilis donors (n = 11) and their transconjugants (P = 0.92).
In the present study, PFGE analysis of 13 E. coli isolates grouped them into 10 clusters, with similarity values ranging from 25% to 100%, indicating high genetic variation among the isolates (Fig. 4A). From Fig. 4A, we observed that isolates from clusters No. 3 and No. 6 did not have exactly the same resistance phenotypes, resistance genes, or virulence genes. This finding revealed that the evolution of resistance and virulence among strains with similar PFGE patterns was not identical, even under the same circumstance. Additionally, isolates such as E21MC5-2, E20MC11-14, E20MC14-4, and E20MC13-5 belonged to different genetic clusters but harbored the same resistance genes and virulence genes.
Figure
4.
The PFGE dendrograms, antimicrobial resistance patterns and related gene distributions among 13 E. coli and 12 P. mirabilis isolates. (A) The dendrogram of E. coli isolates; (B) The dendrogram of P.mirabilis isolates.
The 12 P. mirabilis isolates were grouped into seven clusters, with genetic relatedness ranging from 0% to 100% (Fig. 4B). Similar to E. coli isolates, isolates such as P20MC12-3 and P20MC9-3 from different clusters harbored similar resistance genes. These findings suggested that horizontal gene transfer might exist among these isolates.
4.
Discussion
In this study, we collected 23 samples of dead Crested Ibis embryos from the Mount Emei Biological Resources and Experiment Station Base, Sichuan Province. We identified 13 E. coli (56.52%, 13/23) and 12 P. mirabilis isolates (52.17%, 12/23) from dead ibis embryos, with both bacteria (21.74%, 5/23) simultaneously detected in five samples.
Pathogenicity examination revealed that the mice challenged with the bacteria exhibited symptoms such as depression, anorexia, eye inflammation, and tics, ultimately dying from respiratory failure. The mortality rate in the group challenged by each strain (1×108 CFU per mouse) was 100% within 72 h post-infection, which confirmed the pathogenicity of all strains tested (Takeuchi et al., 2003).
Over the years, AMR is one of the most severe global threats to public health. There is no doubt that overuse of antimicrobials in food animals has expedited the rapid increase and spread of AMR traits and resistant bacteria in the ecosystems (Van et al., 2020; Ardakani et al., 2023). Food animals serve as a major and shared reservoir of resistance determinants for most antimicrobials (Poirel et al., 2018). A study on the AMR trends of 982 animal-derived E. coli samples collected in China revealed an increasing trend in AMR and the presence of antimicrobial resistance genes (ARGs) over the past 50 years (Yang et al., 2022). The correlation analysis also revealed an association between antimicrobial use and the emergence of ARGs at spatial and temporal levels (Yang et al., 2022). In the present study, the results on the antimicrobial resistance in E. coli and P. mirabilis isolates from dead Crested Ibis embryos showed similar resistance patterns: high resistance to amoxicillin (E. coli: 76.92%, P. mirabilis: 91.67%), cefazolin (E. coli: 76.92%, P. mirabilis: 91.67%), oxytetracycline (E. coli: 92.31%, P. mirabilis: 75.00%) and SXT (E. coli: 53.85%, P. mirabilis: 33.33%). More than 30% of isolates were MDR strains, which was consistent with previous surveillance from food animals, particularly the high resistance rates (over 80%) for ampicillin, tetracycline and sulfonamides. The extensive use of these agents in animals likely contributed to the high resistance observed (Poirel et al., 2018; Wu et al., 2018; Girlich et al., 2020). Our survey of antimicrobial use at the Crested Ibis breeding base revealed that antimicrobial administration was strictly controlled and only allowed during infectious disease outbreaks. For instance, amoxicillin and oxytetracycline were used in feed to treat respiratory and intestinal infections, while erythromycin ointment was applied for eye infections. This explains the high resistance to specific antimicrobials (amoxicillin, oxytetracycline) in the present study. Notably, the use of cephalosporins and sulfonamides was rare in the breeding base, yet high resistance to these two drugs (cefazolin, SXT) was observed, implying that resistance genes are widespread in nature and continue to evolve in detrimental ways. Additionally, the horizontal transmission of resistance genes plays a significant role in the acquisition of resistance through mobile genetic elements, in particular plasmids. When this happens, the susceptible bacteria are eliminated by antimicrobial, leaving the resistant strains to replicate through natural selection (Van et al., 2020; Han et al., 2024). Considering the results of the antimicrobial susceptibility test, it is suggested that modern antimicrobials, including the third-generation cephalosporins, aminoglycosides and fluoroquinolones, may be effective in controlling bacterial infections. Meanwhile, biosafety measures should be enhanced, such as cleaning and disinfection of the environment, improving the nutritional status of parent birds, and providing technical training for keepers.
The World Health Organization (WHO) has indicated that the emergence of resistant Enterobacteriaceae bacteria, especially the prevalence of ESBLs-producing strains, poses a serious threat to medicine and public health worldwide in the 21st century (Loayza et al., 2020). Since 2000, CTX-M β-lactamases have increased in prevalence. The global distribution of CTX-Ms is complex; however, blaCTX-M-15 and blaCTX-M-14 are dominant in most countries (Bevan et al., 2017). In China, high diversity of blaCTX-M variants has been observed, with blaCTX-M-55, blaCTX-M-14 and blaCTX-M-65 being the most predominant genotypes in strains from food animals (Rao et al., 2014).
The present study identified similar dominant ESBL genes blaCTX-M-55, blaCTX-M-14 and blaCTX-M-65. The frequency of blaCTX-M-55 (E. coli: 92.31%, P. mirabilis: 50%) was much higher compared to reports from ten years ago (ranging 17.95%–4.30%), but the occurrence of blaCTX-M-65 (E. coli: 15.38%, P. mirabilis: 16.67%) was consistent with these reports (about 18%) (Rao et al., 2014; Tong et al., 2015), indicating that blaCTX-M-55 has become the most common blaCTX-M genotype over time. Correlation analysis revealed a positive correlation between β-lactam resistance tested and the emergence of blaCTX-M genes. Furthermore, the detection rate of blaTEM-1 was very high (100% in E. coli and P. mirabilis) in this study. blaTEM-1 is the first plasmid-mediated β-lactamase resistance gene, and is widespread in Enterobacteriaceae strains from animals (Poirel et al., 2018; Girlich et al., 2020). Although TEM-1 only confers resistance to penicillins and early cephalosporins, its variants’ resistance spectrum has broadened to include advanced cephalosporins, monobactams and β-lactamase inhibitors (Salverda et al., 2010; Girlich et al., 2020). The co-detection rate of blaTEM-1 and blaCTX-M was very high (E. coli: 100%, P. mirabilis: 75%), and most of the resistance genes were transferred through plasmid in conjugation experiments, facilitating high resistance to β-lactams and the widespread spread of resistance.
Antimicrobial resistance and virulence in clinical isolates have been of great concern. In our study, four virulence genes were detected in E. coli: fimC (F1-fimbriae), iucD (aerobactin), iss (serum resistance) and irp2 (iron acquisition system), with iucD and irp2 having the highest prevalence (100%), followed by fimC and iss (92.31%). Previous studies have shown strong positive correlations between the presence of these virulence genes and chicken embryo lethality, with the iucD gene contributing significantly to embryo mortality (Oh et al., 2012). The iucD is involved in iron-chelating ability, and most pathogenic E. coli from poultry express in the aerobactin iron acquisition system, which is strongly associated with virulence (Oh et al., 2012; Hussain et al., 2022). The fimC, iucD and iss are the most common virulence genes found in poultry suffering from colibacillosis, and the presence of irp2 plays an important role in virulence (Hussain et al., 2022). The high prevalence of these virulence genes confirmed that E. coli isolates from dead Crested Ibis embryos were pathogenic, consistent with results of the mouse pathogenicity examination. However, only ucaA (fimbria) with 58.33% prevalence was detected in P. mirabilis, which was not consistent with the high prevalence and multiple virulence genes in isolates from dogs (Liu et al., 2023) and could not fully explain the results of pathogenicity examination in the study. The discrepancy may be due to the selection of a subset of virulence genes for detection based on information from P. mirabilis isolated from ducks (Yu et al., 2020).
Various studies suggested an association between the presence of certain virulence genes and antimicrobial resistance, implying that mechanisms selected for bacterial virulence can also lead to resistance (Sanches et al., 2019; Hussain et al., 2022; Yang et al., 2022). Contrary to previous studies, our study found more common negative correlations between the presence of ESBL genes and virulence genes (except blaCTX-M-14 and iss, r = 0.53). These findings suggest that virulence and antimicrobial resistance may not be independent bacterial characteristics, and the use of antimicrobial agents can lead to increased resistance and positive or negative correlation with virulence factors. The final effect of the correlation between antimicrobial resistance and virulence depends on multiple factors, including the bacterial species, mechanisms of virulence and resistance, host’s immune system and ecological niche (Beceiro et al., 2013).
PFGE serves as a gold standard molecular technique for typing and subtyping many bacteria, including E. coli strains. This genetic analysis can assess the genetic diversity of the entire genome and offer a deeper understanding of the epidemiology and evolution of bacteria (Shin et al., 2014; Vakili et al., 2025). In this study, 13 E. coli and 12 P. mirabilis isolates were analyzed and discriminated using PFGE-Xba I restriction. Dendrogram analysis identified 10 and 7 different PFGE clusters among the 13 E. coli and 12 P. mirabilis isolates, respectively, indicating high genetic variation. This diversity might be explained by the fact that the strains were isolated from dead embryos of different parents and sampled at different times. Additionally, strains with similar PFGE patterns (strains from clusters No. 3 and No. 6 of E. coli, strains from clusters No. 1 and No. 6 of P. mirabilis) showed different resistance phenotypes and genotypes. This suggests that even within similar environments, resistance evolution among clonal strains can vary. Moreover, strains with dissimilar PFGE patterns harbored similar resistance genes (such as E21MC5-2, E20MC11-14, E20MC14-4 and E20MC13-5 in E. coli isolates, P20MC12-3 and P20MC9-3 in P. mirabilis isolates), which suggests that horizontal transfer of resistance gene might exit among these isolates.
5.
Conclusion
According to the current findings of this study, E. coli and P. mirabilis strains were responsible for embryonic mortality in Crested Ibis. Antimicrobial resistance was found to be widespread in this study. A significant number of the strains carried ESBL resistance genes, with blaTEM-1 and blaCTX-M being the most common. Multiple virulence genes (irp2, iucD, fimC and iss in E. coli, ucaA in P. mirabilis) were detected, and the co-presence of ESBL genes and virulence genes was found in many strains. Although there were mainly negative correlations between the presence of the ESBL resistance genes and virulence genes, the horizontal transfer of resistance or virulence genes by mobile genetic elements including plasmids, could produce a large number of resistant and hyper virulent strains, resulting in clinical treatment failure and high mortality. The findings of this study reinforce the need for the strict biosecurity, prudent antimicrobial use and long-term surveillance of bacterial resistance and pathogenicity at the Crested Ibis breeding base.
All animals received humane care according to the criteria outlined in the Guide for the Care and Use of Laboratory Animals. All animal experiments were approved by the Institutional Animal Care and Use Committee of Sichuan Agricultural University (permit No. 20200336).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Aghamohammad, S., Nikbin, V.S., Badmasti, F., Shahcheraghi, F., 2022. High heterogeneity of fecal carriage extended-spectrum beta-lactamase-producing E. coli isolated from iranian community and clinical settings. BMC Infect. Dis. 22, 318. .
Algammal, A.M., Hashem, H.R., Alfifi, K.J., Hetta, H.F., Sheraba, N.S., Ramadan, H., et al., 2021. atpD gene sequencing, multidrug resistance traits, virulence-determinants, and antimicrobial resistance genes of emerging XDR and MDR-Proteus mirabilis. Sci. Rep. 11, 9476. .
Ardakani, Z., Canali, M., Aragrande, M., Tomassone, L., Simoes, M., Balzani, A., et al., 2023. Evaluating the contribution of antimicrobial use in farmed animals to global antimicrobial resistance in humans. One Health 17, 100647. .
Armbruster, C.E., Mobley, H.L.T., Pearson, M.M., 2018. Pathogenesis of Proteus mirabilis infection. EcoSal Plus 8. .
Beceiro, A., Tomás, M., Bou, G., 2013. Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 26, 185-230. .
Bevan, E.R., Jones, A.M., Hawkey, P.M., 2017. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 72, 2145-2155. .
CLSI, 2021. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 31st ed. Clinical and Laboratory Standards Institute, Wayne, PA.
Dallenne, C., Da Costa, A., Decré, D., Favier, C., Arlet, G., 2010. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 65, 490-495. .
El-Tayeb, M.A., Ibrahim, A.S.S., Al-Salamah, A.A., Almaary, K.S., Elbadawi, Y.B., 2017. Prevalence, serotyping and antimicrobials resistance mechanism of Salmonella enterica isolated from clinical and environmental samples in Saudi Arabia. Braz. J. Microbiol. 48, 499-508. .
Elbediwi, M., Tang, Y., Shi, D., Ramadan, H., Xu, Y., Xu, S., et al., 2021. Genomic investigation of antimicrobial-resistant Salmonella enterica isolates from dead chick embryos in China. Front. Microbiol. 12, 684400. .
Fu, C.Z., Guang, X.M., Wan, Q.H., Fang, S.G., 2019. Genome resequencing reveals congenital causes of embryo and nestling death in Crested Ibis (Nipponia nippon). Genome Biol. Evol. 11, 2125-2135. .
Gautom, R.K., 1997. Rapid pulsed-field gel electrophoresis protocol for typing of Escherichia coli O157:H7 and other gram-negative organisms in 1 day. J. Clin. Microbiol. 35, 2977-2980. .
Girlich, D., Bonnin, R.A., Dortet, L., Naas, T., 2020. Genetics of acquired antibiotic resistance genes in Proteus spp. Front. Microbiol. 11, 256. .
Han, Y., Gao, Y.F., Xu, H.T., Li, J.P., Li, C., Song, C.L., et al., 2024. Characterization and risk assessment of novel SXT/R391 integrative and conjugative elements with multidrug resistance in Proteus mirabilis isolated from China, 2018-2020. Microbiol. Spectr. 12, e0120923. .
Hussain, H.I., Iqbal, Z., Iqbal, M., Kuang, X., Wang, Y., Yang, L., et al., 2022. Coexistence of virulence and β-lactamase genes in avian pathogenic Escherichia coli. Microb. Pathog. 163, 105389. .
Karunarathna, R., Ahmed, K.A., Goonewardene, K., Gunawardana, T., Kurukulasuriya, S., Liu, M., et al., 2022. Exposure of embryonating eggs to Enterococcus faecalis and Escherichia coli potentiates E. coli pathogenicity and increases mortality of neonatal chickens. Poult. Sci. 101, 101983. .
Kojima, A., Ishii, Y., Ishihara, K., Esaki, H., Asai, T., Oda, C., et al., 2005. Extended-spectrum-beta-lactamase-producing Escherichia coli strains isolated from farm animals from 1999 to 2002: report from the Japanese veterinary antimicrobial resistance monitoring program. Antimicrob. Agents Chemother. 49, 3533-3537. .
Liu, L., Dong, Z., Ai, S., Chen, S., Dong, M., Li, Q., et al., 2023. Virulence-related factors and antimicrobial resistance in Proteus mirabilis isolated from domestic and stray dogs. Front. Microbiol. 14, 1141418. .
Loayza, F., Graham, J.P., Trueba, G., 2020. Factors obscuring the role of E. coli from domestic animals in the global antimicrobial resistance crisis: an evidence-based review. Int. J. Environ. Res. Publ. Health 17. .
Lu, J., Yang, Y., Wu, Y., Liu, C., Zeng, Y., Lei, L., et al., 2022. Escherichia coli carrying IncI2 plasmid-mediated mcr-1 genes in crested ibis (Nipponia nippon). J. Infect. Public Health 15, 558-561. .
Meng, Q., Wang, S., Han, X., Han, Y., Ding, C., Dai, J., et al., 2014. Multiplex PCR assay for detection of virulence genes in avian pathogenic Escherichia coli. Acta Microbiol. Sin. 54, 696-702. .
Mirzaei, A., Nasr Esfahani, B., Raz, A., Ghanadian, M., Moghim, S., 2021. From the urinary catheter to the prevalence of three classes of integrons, β-lactamase genes, and differences in antimicrobial susceptibility of Proteus mirabilis and clonal relatedness with Rep-PCR. BioMed Res. Int. 2021, 9952769. .
Oh, J.Y., Kang, M.S., Yoon, H., Choi, H.W., An, B.K., Shin, E.G., et al., 2012. The embryo lethality of Escherichia coli isolates and its relationship to the presence of virulence-associated genes. Poult. Sci. 91, 370-375. .
Paixão, A.C., Ferreira, A.C., Fontes, M., Themudo, P., Albuquerque, T., Soares, M.C., et al., 2016. Detection of virulence-associated genes in pathogenic and commensal avian Escherichia coli isolates. Poult. Sci. 95, 1646-1652. .
Poirel, L., Madec, J.Y., Lupo, A., Schink, A.K., Kieffer, N., Nordmann, P., et al., 2018. Antimicrobial resistance in Escherichia coli. Microbiol. Spectr. 6. .
Rao, L., Lv, L., Zeng, Z., Chen, S., He, D., Chen, X., et al., 2014. Increasing prevalence of extended-spectrum cephalosporin-resistant Escherichia coli in food animals and the diversity of CTX-M genotypes during 2003-2012. Vet. Microbiol. 172, 534-541. .
Rezaee, M.S., Liebhart, D., Hess, C., Hess, M., Paudel, S., 2021. Bacterial infection in chicken embryos and consequences of yolk sac constitution for embryo survival. Vet. Pathol. 58, 71-79. .
Salverda, M.L., De Visser, J.A., Barlow, M., 2010. Natural evolution of TEM-1 β-lactamase: experimental reconstruction and clinical relevance. FEMS Microbiol. Rev. 34, 1015-1036. .
Sanches, M.S., Baptista, A.A.S., de Souza, M., Menck-Costa, M.F., Koga, V.L., Kobayashi, R.K.T., et al., 2019. Genotypic and phenotypic profiles of virulence factors and antimicrobial resistance of Proteus mirabilis isolated from chicken carcasses: potential zoonotic risk. Braz. J. Microbiol. 50, 685-694. .
Shin, S.W., Byun, J.W., Jung, M., Shin, M.K., Yoo, H.S., 2014. Antimicrobial resistance, virulence genes and PFGE-profiling of Escherichia coli isolates from South Korean cattle farms. J. Microbiol. 52, 785-793. .
Takeuchi, K., Smith, M.A., Doyle, M.P., 2003. Pathogenicity of food and clinical Listeria monocytogenes isolates in a mouse bioassay. J. Food Protect. 66, 2362-2366. .
Tong, P., Sun, Y., Ji, X., Du, X., Guo, X., Liu, J., et al., 2015. Characterization of antimicrobial resistance and extended-spectrum β-lactamase genes in Escherichia coli isolated from chickens. Foodb. Pathog. Dis. 12, 345-352. .
Tran, T., Checkley, S., Caffrey, N., Mainali, C., Gow, S., Agunos, A., et al., 2021. Genetic characterization of AmpC and extended-spectrum beta-lactamase phenotypes in Escherichia coli and Salmonella from Alberta broiler chickens. Front. Cell. Infect. Microbiol. 11, 622195. .
Vakili, M., Goli, H., Javidnia, J., Alipour, T., Eslami, M., 2025. Genetic diversity and antibiotic resistance patterns of Escherichia coli isolates causing septicemia: a phylogenetic typing and PFGE analysis. Diagn. Microbiol. Infect. Dis. 111, 116586. .
Van, T.T.H., Yidana, Z., Smooker, P.M., Coloe, P.J., 2020. Antibiotic use in food animals worldwide, with a focus on Africa: pluses and minuses. J. Glob. Antimicrob. Resist. 20, 170-177. .
Wu, C., Wang, Y., Shi, X., Wang, S., Ren, H., Shen, Z., et al., 2018. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008-2014. Emerg. Microb. Infect. 7, 30. .
Xi, Y., Lu, B., Fujihara, N., 2001. Captive rearing and breeding of the crested ibis, Nipponia nippon. J. Poultry Sci. 38, 213-224. .
Xi, Y., Wood, C., Lu, B., Zhang, Y., 2007. Prevalence of a septicemia disease in the crested ibis (Nipponia nippon) in China. Avian Dis. 51, 614-617. .
Yang, H.Q., Chen, X.L., Li, Y.D., 2020. Analysis of obstacles and causes encountered in artificial breeding of Crested ibis in Sichuan. J. Sichuan For. Sci. Technol. 41, 84-89. (in Chinese).
Yang, L., Shen, Y., Jiang, J., Wang, X., Shao, D., Lam, M.M.C., et al., 2022. Distinct increase in antimicrobial resistance genes among Escherichia coli during 50 years of antimicrobial use in livestock production in China. Nat. Food 3, 197-205. .
Yu, Z., Yang, K., Wang, Y., Zhang, Q., 2020. Isolation and identification of Proteus mirabilis from Muscovy ducks and analysis of its virulence genes. Chin. J. Anim. Infect. Dis. 28, 14-20. (in Chinese).


 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds