Passerine mimics often imitate various vocalizations from other bird species and incorporate these sounds into their song repertoires. While a few anecdotes reported that wild songbirds imitated human-associated sounds, besides captive parrots and songbirds, systemic and quantitative studies on human-made sound mimicry in wild birds remain scarce. In this study, we investigated the mimetic accuracy and consistency of electric moped sounds imitated by an urban bird, the Chinese Blackbird (Turdus mandarinus). We found that: (1) Only one type of electric moped sound was imitated, i.e., 13 of 26 males mimicked the first part of the antitheft alarm, a phrase containing a series of identical notes. (2) The mimicry produced by male Chinese Blackbirds had fewer notes and lower consistency within phrases compared to the model alarms. (3) The mimicry of male Chinese Blackbirds was imperfect, i.e., most of the acoustic parameters differed from the model alarms. Additionally, mimetic notes were lower in frequency than the models. Mimetic notes from two areas were also different in acoustic structures, suggesting Chinese Blackbirds might learn mimicry mainly from conspecific neighbors within each area respectively rather than electric mopeds, namely the secondary mimicry. Imperfect mimicry of human-made sounds could result from cost and physical constraints, associated with high consistency, frequency, and repetitions. Consequently, Chinese Blackbirds copied a simplified version of electric moped alarms. We recommend further attention to mimic species inhabiting urban ecosystems to better understand vocal mimicry's adaptation to ongoing urbanization.
Many passerines imitate various vocalizations from other bird species, and incorporate these mimetic sounds into their song repertoires (Dalziell et al., 2015; Dalziell and Welbergen, 2016), including Superb Lyrebird (Menura novaehollandiae) (Dalziell et al., 2021), Northern Mockingbird (Mimus polyglottos) (Gammon and Corsiglia, 2019), and Icterine Warbler (Hippolais icterina) (Jůzlová and Riegert, 2012). Although most mimetic sounds were found imitated from other bird species (Gammon and Altizer, 2011; Kelley and Healy, 2011; Jůzlová and Riegert, 2012), some passerines also mimic sounds from non-avian species. For instance, Northern Mockingbirds imitate calls from at least 12 anuran species across North America (Gammon and Corsiglia, 2019); Long-tailed Shrikes (Lanius schach) imitate the calling songs of the male Cicada [Meimuna mongolica (Distant)] in China (http://www.xeno-canto.org/839067). Some mimic species are even reported to imitate the sounds of inanimate objects, such as human-made sounds and artificial noises (Dalziell et al., 2015). Most reports of human-made sound mimicry were found in captive songbirds, parrots, and even non-passerines (ten Cate and Fullagar, 2021; Benedict et al., 2022). For instance, a captive Australian Musk Duck (Biziura lobata) imitated the sound of closing a metal aviary door (ten Cate and Fullagar, 2021), while many pet parrot species imitated human words (Benedict et al., 2022). Human-made sounds imitated by wild birds have often been reported as casual anecdotes, but to our knowledge, a systematic and quantitative analysis has never been attempted by any scientific study. The only published example of which we are aware of a wild bird imitating a human-made sound is European Starlings (Sturnus vulgaris) from New Zealand mimicking the shepherd's whistle (see Fig. 4 in Hausberger et al., 1991). Even for this example, we lack a description and spectrogram of the model sound.
One reason why human-made sound mimicry is understudied is that some of the best-studied mimics, such as Albert's Lyrebird (Menuraalberti) (Pavlacky et al., 2015; Backhouse et al., 2022) and Satin Bowerbirds (Ptilonorhynchus violaceus) (Coleman et al., 2007), inhabit forests in which exposure to human sounds is very low. These wild birds are less exposed to sounds from humans, and thus less mimic human-made sounds. To conduct such studies, it is more appropriate to choose a species living close to human society, namely, the urban areas. Under these scenarios, the Chinese Blackbird (Turdus mandarinus), a songbird producing elaborate vocal mimicry (Fu et al., 2024), provides us with an ideal opportunity to study human-made sound mimicry in wild birds. Chinese Blackbird is one of the most common bird species inhabiting urban environments in cities of southeast China (Zhao et al., 2023; Zheng, 2023), and they are exposed to various human-associated sounds, which may be the potential putative models to be imitated. As transport mode has been largely developed, electric mopeds are one of the most prevalent transportation modes in China (Sun and Wang, 2023): by 2022, over 350 million electric bikes/mopeds were registered in China (National Bureau of Statistics, 2023). Electric mopeds often produce a series of identical sounds with high consistency, such as alarms for antitheft, and these sounds are potentially possible to be imitated by sympatric Chinese Blackbirds.
Vocal consistency in songbirds is an honest signal to assess males’ quality: high-quality males produce highly consistent songs, which are difficult to produce (Sakata and Vehrencamp, 2012). Female Blue Tits (Cyanistes caeruleus), for instance, prefer males with high levels of consistent repetition within songs (Sierro et al., 2023). To repeat identical syllables within a song phrase with high consistency, males need to accurately coordinate their syringeal muscles, breaths, and neural control (Lambrechts and Dhondt, 1988; Zollinger and Suthers, 2004; Sakata and Vehrencamp, 2012; Sun et al., 2022b). Consequently, highly consistent songs are costly to produce. However, it has not yet been investigated whether an avian mimic can accurately imitate the high consistency of artificial sounds or not. Additionally, mimics usually imitate a short version or less repetition of a vocalization, compared to models (Dalziell and Magrath, 2012; Fu et al., 2024). For example, superb lyrebirds produce fewer repetitions of identical elements within songs than the models, the songs of the Grey Shrike-thrush (Colluricincla harmonica) (Dalziell and Magrath, 2012). However, it also remains to know if this is also the case in human-made sound mimicry, namely, less notes within a phrase in mimicry.
In this study, we investigated human-made sound mimicry by analyzing the mimicry of electric moped sounds by Chinese Blackbirds. Specifically, we aimed to address three questions. First, do Chinese Blackbirds mimic the sounds of electric mopeds? If so, what type of sounds are mimicked? Second, do Chinese Blackbirds imitate these sounds with accurate consistency? We hypothesized that blackbirds' mimicry is lower in consistency compared to the model sounds, as producing highly consistent sounds is inherently challenging (Sakata and Vehrencamp, 2012). Third, we evaluated the mimetic accuracy between the model and mimetic sounds through systemic acoustic parameter analyses, including the number of notes per song phrase. We predicted that the blackbirds can produce imperfect mimicry of electric moped sounds, given the difficulty of imitating human-made sounds and the distinctive characteristics of blackbirds’ vocal mimicry (Fu et al., 2024).
2.
Materials and methods
2.1
Study area and species
The Chinese Blackbird (Turdus mandarinus) is a common resident thrush (weight range: 80–125 g) inhabiting the urban environment of Nanjing, China (Fig. 1A and B) (del Hoyo et al., 2020; Zhao et al., 2023; Zheng, 2023; Liu and Li, 2024). The males are highly territorial during breeding season and produce large song repertoires (Fu et al., 2024). They produce lots of songs during the breeding season in high perches from trees or buildings (approximately 5–20 m above ground), singing with immediate variety, and each song contains several phrases, including both various mimetic units and species-specific song elements (Fu et al., 2024). We studied electric moped sound mimicry of blackbirds in Xianlin campus of Nanjing University (area 1, 32.12°N, 118.95°E) and an adjacent residential area (area 2, 32.13° N, 118.98° E, Fig. 1C). Thousands of electric mopeds are used in these areas with dense human population, producing loud noise including horns and alarms, which can be potential models for Chinese Blackbirds to mimic. We recorded song repertoires of Chinese Blackbirds and electric moped sounds in the above study areas during the breeding season (February–August 2023).
Figure
1.
The maps showing the location of the study areas (1 and 2) and territories of Chinese Blackbirds. Maps of (A) China and (B) Nanjing were generated using the ArcGIS 10.8 (http://desktop.arcgis.com/cn/) based on the standard map GS (2024)0650 downloaded from National Platform for Common GeoSpatial Information Services (https://cloudcenter.tianditu.gov.cn/), without modifications of the boundaries. (C) A map showing the territories of 26 male Chinese Blackbirds whose song repertoires were recorded, which was generated using the Google Earth Pro 7.3 (https://earth.google.com). Individuals from area 1 and area 2 are indicated by red and orange dashed circles respectively. Of 26 males, 13 imitated the alarm of the electric moped (dots in light blue), while the remaining 13 males did not (dots in yellow).
Although we did not use color banding to identify individual males, we identified the territories of male Chinese Blackbirds by observation. Males usually sing in several specific perches from trees or the roofs of buildings, and the connecting lines between these perches compose the border of each territory (Fu et al., 2024). Males would chase their neighbors (i.e. other males) who strayed into their territory, which is enough to discriminate focal individuals and their neighbors. Territories of 26 males were identified in total (Fig. 1C). Song repertoires of males were recorded using a handheld RODE NTG4 + directional microphone and a TASCAM Portacapture X8 portable recorder (TEAC Co., Ltd, Tokyo) with 48 kHz sample rate and 16/24-bit resolution. All recordings were saved as WAV format. Weather without strong wind during daytime (8:00–19:00 h) was required for field work. Recordings were made with 10–20 m to focal males. We recorded >498 min of song repertoires from 26 males with a mean duration of 19.165 ± 11.741 min (Mean ± SD, N = 26, 2.783–63.100 min) per male, including a total of 3041 songs (117 ± 53 songs per male, N = 26, 14–220 songs).
2.3
Recording putative model sounds
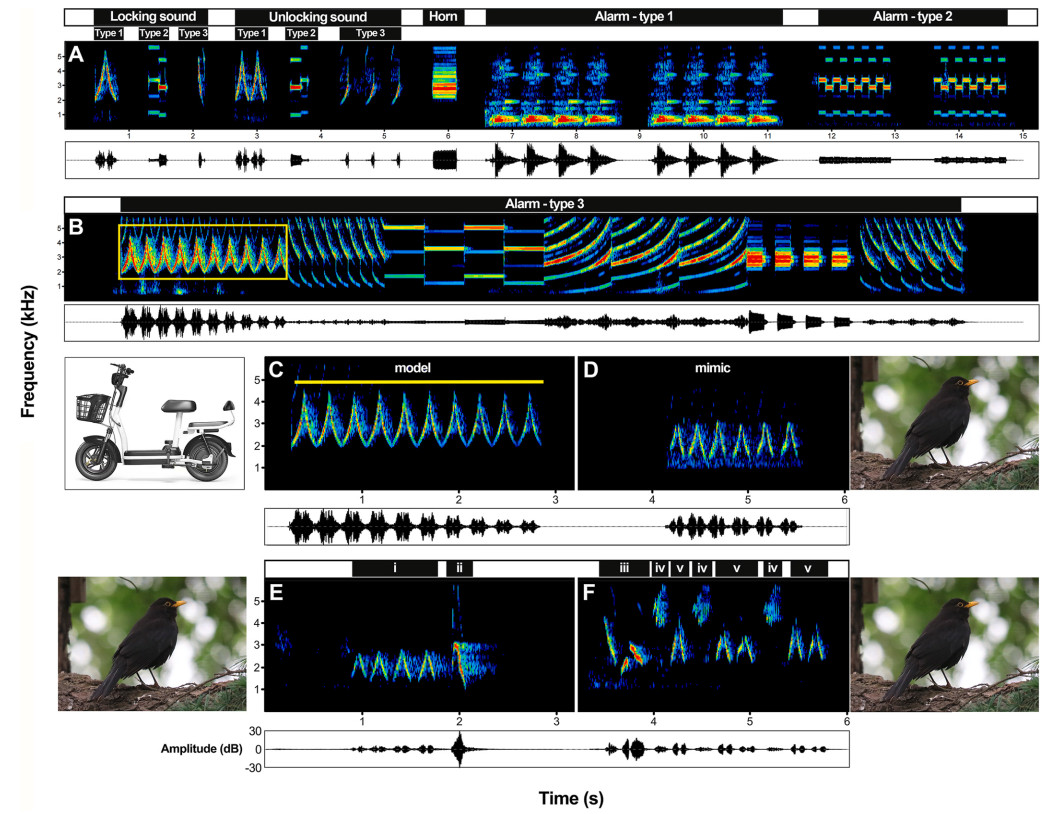
Sounds of electric mopeds include four categories: (1) locking sounds, a prompt tone indicates that the electric moped is locked by key; (2) unlocking sounds, a prompt tone indicates that the electric moped is unlocked by key; (3) horn, a sound to alert pedestrians; (4) antitheft alarm (hereafter alarm), a warning tone produces when an electric moped is moved without unlocking. Electric mopeds of different types or brands have various above sounds (e.g. Fig. 2A and B, Appendix Audio S1). For example, one brand of moped produces a type 1 locking sound, a type 1 unlocking sound, and a type 3 alarm (Fig. 2A–B, Appendix Audio S1); another brand produces a type 2 locking sound, a type 2 unlocking sound, and a type 2 alarm (Fig. 2A–Appendix Audio S1).
Figure
2.
Example spectrograms showing the electric moped sounds and the imitations produced by Chinese Blackbirds (related to Appendix Audio S1). (A)–(B) Three examples of locking sound, unlocking sound, and alarms from three brands of electric mopeds respectively, and one example showing the horn. (C) The first part of type 3 alarm (containing 10 continuous notes), which is the model sound mimicked by Chinese Blackbirds. (D) A mimetic alarm produced by a Chinese Blackbird, containing six continuous notes. (E) A mimetic song containing mimetic alarm; ⅰ: a mimetic phrase containing four continuous notes; ⅱ: a mimetic solo of Masked Laughingthrush (Pterorhinus perspicillatus). (F) A mimetic song containing mimetic alarms/locking/unlocking sounds, which are interspersed by other song phrases; ⅲ: a species-specific song phrase of Chinese Blackbirds; ⅳ: mimetic calls of White-cheeked Starling (Spodiopsar cineraceus); ⅴ: mimetic alarms of electric moped.
To record model sounds, we first contacted the owners of electric mopeds for help, because locking/unlocking sounds could be only induced by the keys held by owners. The alarms were induced by shaking the mopeds. We recorded the sounds from over 40 electric mopeds around the territories of the 26 blackbirds to cover putative model sounds from multiple brands as much as possible. We recorded 1–4 sound types from each moped because some mopeds lacked some of the above sound types (1)–(3) or only had alarms, and each sound type was recorded 1–2 times. All mopeds were stationary and recordings were made with 10–20 m to them.
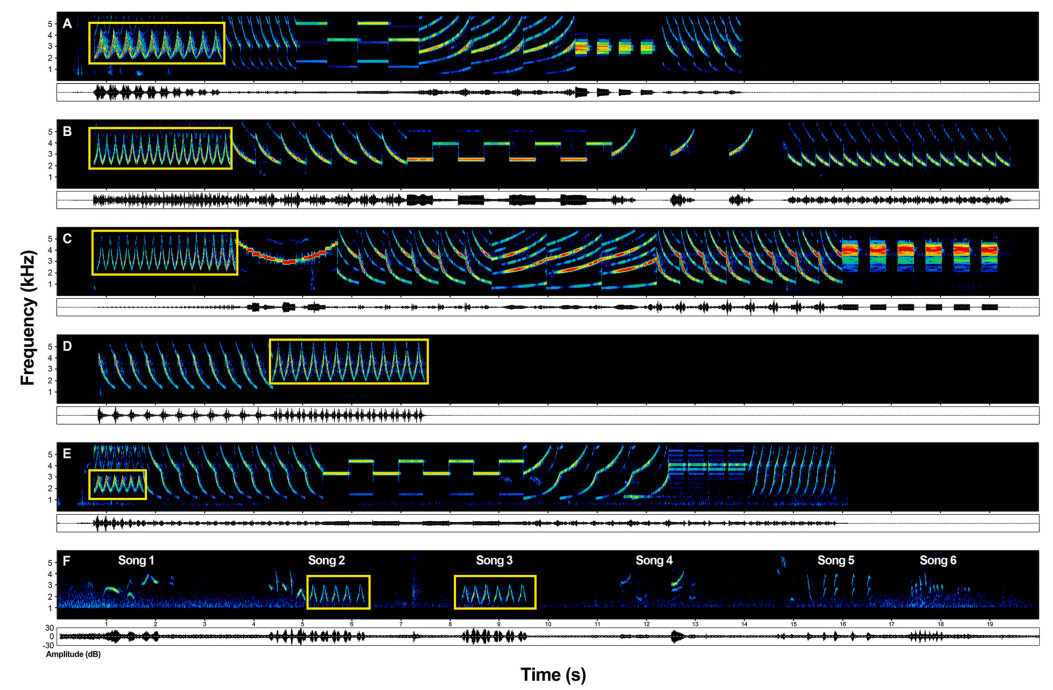
Electric mopeds of different brands sometimes produce identical or similar sounds. For example, one of the most common alarms contains a phrase consisting of a series of identical notes (each note includes a rising and a falling part, Figs. 2B and 3A-E, Appendix Audio S2). This phrase is used by multiple brands, with variations in frequency or duration. It is impossible to decide which brand the phrase is imitated from if the Chinese Blackbirds mimic this phrase inaccurately. Consequently, we recorded the alarms containing similar phrases from mopeds of different brands as much as possible, which were used for subsequent analyses (such as Fig. 3A–E).
Figure
3.
Example spectrograms showing electric moped alarms from five brands (A)–(E) (related to Appendix Audio S2), which contained similar phrases (indicated by bright yellow boxes), and six songs of Chinese Blackbirds (F) (related to Appendix Audio S3). Two songs (2 and 3) in (F) contained a mimetic alarm phrase.
We checked all recordings of blackbird songs to find mimetic electric moped sounds using Avisoft-SASLab Lite (https://www.avisoft.com/) with settings: Fast Fourier Transform (FFT) 256, FlatTop window, frame size 100%, bandwidth 235 Hz, resolution 63 Hz, overlap 50%, temporal resolution 8 ms. In this study, we identified phrase as the sound composed by a series of identical notes (such as Fig. 2C and D, Appendix Audio S1), since sounds of electric mopeds are usually stereotyped. All mimetic phrases were identified through spectrogram comparisons and recording listening by two researchers respectively (C.F. and X.W.). Each identified mimetic phrase should be agreed by both of the researchers. Many mimic species, including Chinese Blackbirds, extract notes from different models and combine these mimetic sounds into one single song (Gammon and Altizer, 2011; Dalziell et al., 2015; Fu et al., 2024). If a song contained mimetic sounds of electric mopeds (e.g., Figs. 2E and 3F, Appendix Audio S3), we clipped the song into a single recording for subsequent analyses. Locking/unlocking sounds sometimes possess identical or similar structures to alarms (e.g., type 1 locking/unlocking sounds and the first part of the type 3 alarm, Fig. 2A–B). To decide what sounds of mopeds were imitated by the Chinese Blackbirds, we conducted the following procedure.
(1) If a mimetic phrase contained ≥3 notes, we identified it as a mimetic alarm.
(2) If a mimetic phrase contained one or two notes with distinct structures to alarms, we identified it as a mimetic locking/unlocking sound.
(3) If a mimetic phrase contained one or two notes with similar structures to alarms, we identified it as a mimetic alarm/locking/unlocking sound.
2.5
Evaluating consistency
Only sounds with low background noise (<1 kHz) were selected for subsequent analyses, and we prepared all recordings with high-pass filtered at 1 kHz and low-pass filtered at 6 kHz, to reduce the background noise. We used similarity coefficients to evaluate the vocal consistency for both model and mimetic electric moped sounds, which were calculated using the tool batch correlator in Raven Pro 1.6.5 (K. Lisa Yang Center for Conservation Bioacoustics, 2024), with the following settings: FFT 512, Hamming window, 90% overlap. Similarity coefficient is spectrogram similarity between continuous notes within a phrase, ranging from zero to one: zero indicates no similarity between two notes, while one indicates identical (de Kort et al., 2009; Sprau et al., 2013; Sun et al., 2022b). The similarity coefficients were calculated by comparing each note with all other notes within a phrase using a spectrogram cross-correlation algorithm (Cortopassi and Bradbury, 2000; Cramer, 2012; Sierro et al., 2022), which measures the maximum pixel-by-pixel similarity between two spectrograms (Cramer, 2013). Then the coefficients of all comparisons were averaged, and each phrase thus had one single coefficient (Sun et al., 2022b). To calculate the coefficients, we chose 1–3 phrases per blackbird and electric moped with high-quality (high signal-to-noise ratio) respectively, and chose 3–7 continuous notes per phrase (each note was clipped into one single recording).
2.6
Acoustic analysis
To evaluate mimicry accuracy, nine acoustic parameters were measured for each note.
(1) Duration (D, s): the time length of a note.
(2) Duration 90% (D90, s): the duration including 90% of the energy in a note.
(3) High frequency (Fhigh, Hz): the maximum frequency of a note.
(4) Low frequency (Flow, Hz): the minimum frequency of a note.
(5) Peak frequency (Fpeak, Hz): the frequency at which maximum energy occurs.
(6) Frequency range (Frange, Hz): the frequency difference between a note's maximum and minimum frequency.
(7) Bandwidth 90% (B90, Hz): the frequency bandwidth within which 90% of the note's energy falls.
(8) Frequency 5% (F5, Hz): the frequency below which 5% of the note's energy lies.
(9) Frequency 95% (F95, Hz): the frequency above which 5% of the note's energy lies.
Additionally, the number of notes each phrase contained was also counted. We did not include frequency modulation (FM) in our analysis, but since we included duration, number of notes, and bandwidth, aspects of FM are represented in the data.
2.7
Statistical analysis
To compare the number of notes per phrase between model and mimetic sounds, a Kruskal–Wallis rank sum test (KW test) was performed using kruskal. test function in R (R Core Team, 2023). We also conducted a KW test to investigate the difference of similarity coefficients between model and mimetic sounds. To evaluate mimicry accuracy, we conducted three analyses.
(1) We first conducted a hierarchical clustering analysis (HCA) based on nine acoustic parameters using hclust function in R package ‘stats’, and dendrogram was drawn using ggtree function based on R packages ‘‘ggplot2’, ‘ggtree’, ‘treeio’, ‘ggdendro’, and ‘ape’ (Yu et al., 2017; Paradis and Schliep, 2019; Wang et al., 2020).
(2) We conducted the principal component analysis (PCA) to condense nine parameters using princomp function in R package ‘psych’ (Revelle, 2024). Before the PCA, we performed the Kaiser–Meyer–Olkin (KMO) and Bartlett's test of sphericity to test if our data are suitable for conducting PCA. The overall sampling adequacy measure was >0.6 and all p values of Bartlett's test were <0.001: both of the results indicated that our data were acceptable for the PCA (Budaev, 2010). We extracted PC1 and PC2, and performed KW tests to compare the PC1 and PC2 between model and mimetic sounds.
(3) We conducted Shapiro–Wilk tests and Levene's tests for each parameter, and all parameters did not obey normal distribution. KW tests were thus conducted to compare differences between model and mimetic sounds for each parameter.
All statistical analyses were carried out using R 4.4.1 in the RStudio 2024.04.2 + 764. All p values were two-tailed, and statistical significance was set as p < 0.05.
3.
Results
3.1
Overview
According to our investigation, 13 from 26 male Chinese Blackbirds (50%) mimicked the alarm of electric mopeds (Fig. 1C). We identified 84 songs contained mimetic alarms from 3041 songs (2.8%) in total. Chinese Blackbirds only mimicked the first part of type 3 alarm of electric mopeds (Fig. 2D), containing a series of continuous identical notes (Fig. 2C and D, Appendix Audio S1). From the 13 individuals, nine (69%, individuals 2–7, 11, 16, and 20, Fig. 1C) produced phrases containing a series of continuous notes of mimetic alarm (Fig. 2D), or incorporated these phrases into one song (Fig. 2E–Appendix Audio S1). The remaining four individuals (4/13, 31%, individuals 22, 23, 25 and 26, Fig. 1C) interspersed the mimetic alarm (or locking/unlocking sounds) with the species-specific songs or other mimetic sounds (Fig. 2F–Appendix Audio S1).
3.2
The number of notes and consistency
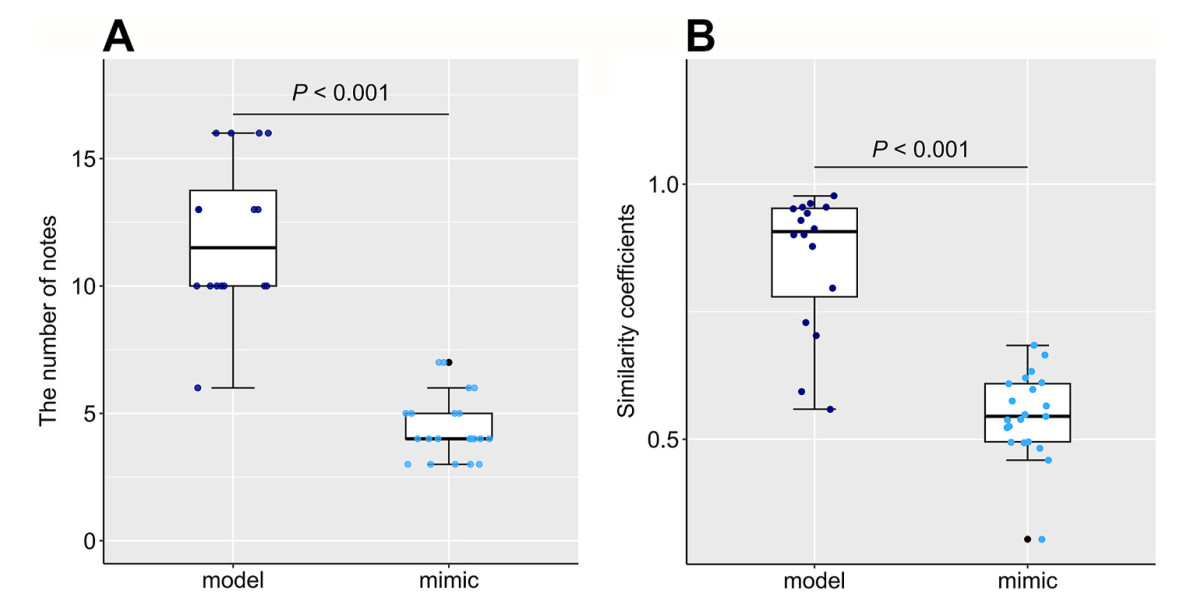
To compare the number of notes between the model and mimetic phrases, we used 21 mimetic phrases from nine Chinese Blackbirds and 16 phrases from 13 electric mopeds. Chinese Blackbirds imitated fewer notes per phrase compared to real alarm of electric mopeds (H = 26.193, df = 1, p < 0.001; Fig. 4A). Mimetic phrases contained 3–7 continuous notes (mean ± SD = 4.429 ± 1.248, N = 9, np = 21, N indicates the number of individuals, while np indicates the number of phrases), while model alarms contained 6–16 continuous notes (12 ± 2.966, N = 13, np = 16).
Figure
4.
Boxplots and dots showing the differences between model and mimetic alarms for (A) the number of notes per phrase and (B) the vocal consistency (i.e. similarity coefficients). The thick lines of the boxplots indicate the medians, the upper and lower lines of boxplots indicate the 25% and 75% quartiles. The whiskers encompass outermost data within 1.5 times the interquartile ranges. The colored dots present raw observations, and the black dots present outlying data points (data beyond the end of the whiskers).
To compare vocal consistency, we used a total of 21 mimetic phrases containing 93 notes from nine Chinese Blackbirds, and 16 phrases containing 83 notes from 13 electric mopeds. The consistency of notes within mimetic phrases (measured as similarity coefficients) was significantly lower than the consistency of notes of the moped alarm sounds (H = 21.719, df = 1, p < 0.001; Fig. 4B). Similarity coefficients of models were 0.559–0.977 (0.853 ± 0.136, N = 13, np = 16), while mimetic alarms were 0.304–0.684 (0.548 ± 0.083, N = 9, np = 21).
3.3
Mimicry accuracy
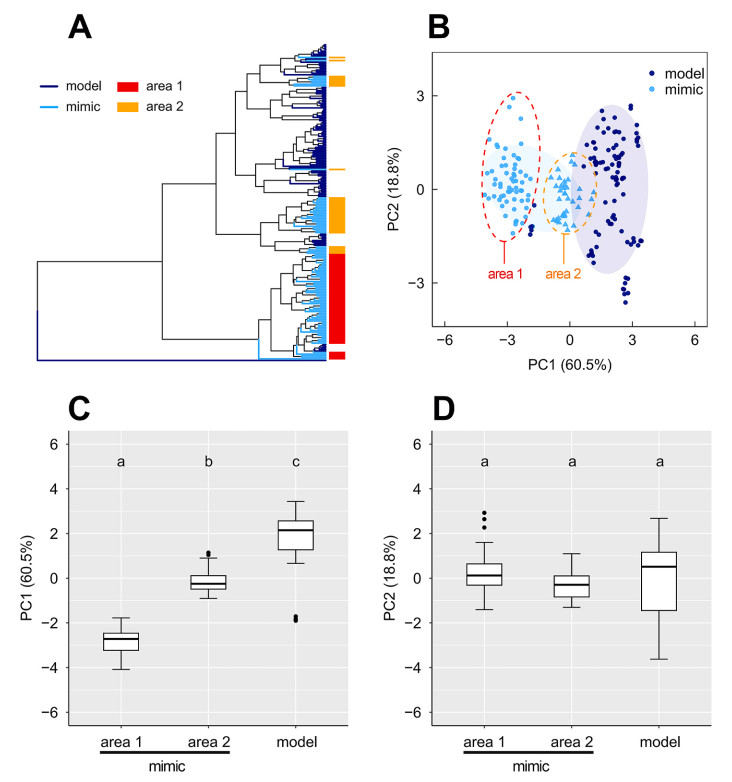
To evaluate mimicry accuracy, 100 mimetic notes (N = 13) and 101 model notes (N = 13) of alarms were extracted and measured. According to HCA and PCA, most mimetic and model notes were discriminated clearly (Fig. 5A and B). PC1 (frequency parameters contributed mostly, including Fhigh and F95) and PC2 (temporal parameters contributed mostly, including D and D90) together explained 79.3% of the variance (Table 1). PC1 of model was higher than the mimetic notes (H = 153.348, df = 2, p < 0.001; Fig. 5C), i.e., mimetic notes were generally lower pitched than the model, while PC2 was not different significantly (H = 5.589, df = 2, p = 0.061; Fig. 5D). Mimetic notes from areas 1 and 2 (Fig. 1C) were also classified clearly by HCA and PCA (Fig. 5A–B): the mimetic alarms from area 1 were lower in frequencies than those from area 2 (p < 0.001, Fig. 5C).
Figure
5.
Results of mimicry accuracy. (A) Dendrogram shows the results of hierarchical clustering analysis based on nine acoustic parameters. Mimetic notes from area 1 and area 2 are indicated by red and orange bars respectively. (B) Scatter diagram showing results of principal component analyses classifying model and mimetic alarms. Mimetic notes from area 1 and area 2 are indicated by red and orange dashed circles respectively. (C) Comparisons of PC1 (Fhigh and F95 contributed mostly) between model and mimetic alarms. (D) Comparisons of PC2 (D and D90 contributed mostly) between model and mimetic alarms. Statistical significance is indicated by letters. The thick lines of the boxplots indicate the median, the upper and lower lines of boxplots indicate the 25% and 75% quartiles. The whiskers encompass outermost data within 1.5 times the interquartile ranges, and the black dots present outlying data points (data beyond the end of the whiskers).
Table
1.
The factor loadings of principal component analysis for nine acoustic parameters of model and mimetic alarms. Loadings >0.40 are shown in bold.
All acoustic parameters differed significantly between mimetic and model notes except D90 (Table 2). Duration of mimetic notes were shorter than the models (p < 0.001; Table 2). Frequency parameters, including Fhigh, Flow, Fpeak, F5, and F95 of mimetic notes were lower than the models (p < 0.001; Table 2). Frange of mimetic notes was narrower than the models (p < 0.001), but B90 of mimetic notes was broader than the models (p < 0.001; Table 2).
Table
2.
Mean (SD) and differences between model and mimetic alarms for each acoustic parameter. N indicates the number of individuals; np indicates the number of phrases; nn indicates the number of notes.
In this study, we found that Chinese Blackbirds could imitate a human-made sound, the antitheft alarm of electric mopeds. However, they possessed imperfect mimicry. As prior studies only reported anecdotal evidence (Hausberger et al., 1991; Dalziell et al., 2015), our study provided quantitative and sympatric investigation and demonstrated that a passerine mimic can imitate highly consistent artificial sounds with special features. Human-made sound imitations also reflect the adaptations of urban birds in vocal mimicry to human-dominated environments.
We found that male blackbirds did not accurately imitate the consistency, the number of notes, and acoustic parameters of model sounds. There may be two explanations for the imperfect mimicry of human-made sounds: (1) energy saving (Fu et al., 2024), and (2) physical constraints (Zollinger and Suthers, 2004; Zann and Dunstan, 2008), or the combination of them (Dalziell et al., 2015). Many model sounds, such as electric moped alarms, were costly to produce, or beyond the ability of the mimics to imitate such sounds with high consistency, long duration, or broad frequency range (i.e. high vocal performance) (Zann and Dunstan, 2008; Sakata and Vehrencamp, 2012). Consequently, producing a simplified imitation of model sounds has been found across some bird mimics (Zann and Dunstan, 2008; Dalziell and Magrath, 2012). For instance, Chinese Blackbirds produce shorter mimetic cuckoo calls than the models, though these cuckoo calls are simple in acoustic structures (Fu et al., 2024). Superb Lyrebirds copy a less repetitive version of Grey Shrike-thrush songs (Dalziell and Magrath, 2012), and also cannot match the broad bandwidth of whipcrack element of the Eastern Whipbird (Psophodes olivaceus) songs (Zann and Dunstan, 2008). Systematic analysis in non-avian sound mimicry is rarely conducted, but a descriptive investigation found that Northern Mockingbirds seem to copy less harmonic versions of anuran calls (Gammon and Corsiglia, 2019). Vocal consistency has not yet to be investigated systematically in vocal mimicry. However, a lower consistency in blackbird mimicry than sounds of electric mopeds also suggested energy saving or constraints, considering highly consistent vocalizations were hard to produce (Sakata and Vehrencamp, 2012). In line with broad bandwidth and fast trill rate (vocal deviation) in birdsongs (Phillips and Derryberry, 2017a, 2017b), high vocal consistency also serves high vocal performance, namely, high similarity among a series of identical notes within a song phrase, which is costly and hard to produce, and further indicates the high quality of males (Sakata and Vehrencamp, 2012; Sierro et al., 2023). Our results revealed that Chinese Blackbirds mimicked human-made sounds through a combination of simplified strategies, including reducing consistency, note repetitions, and frequencies.
However, D90 did not differ between model and mimetic notes, and B90 of mimetic notes were even broader than the models. We thus argue that Chinese Blackbirds might try to resemble the acoustic structure of antitheft alarms of electric mopeds as much as possible, though they had to transform the mimicry because of cost and constraints. Electric moped alarms obviously possess no ecological function to Chinese Blackbirds’ life history. Thus the mimicry of the alarms can be the result of promoting versatility, i.e., the ability to mimic many different sounds (Dalziell et al., 2015). We suggest that human-made sound mimicry might be evidence to support the sexual selection hypothesis in vocal mimicry of Chinese Blackbirds, i.e., mimicry is a signal of male quality (Dalziell et al., 2015). A few studies have proved the sexual selection hypothesis in vocal mimicry, including the handicap hypothesis, (accuracy and versatility) (Coleman et al., 2007), and the sensory trap hypothesis (Dalziell et al., 2021). For example, the number of model species mimicked by the male Satin Bowerbirds was positively related to mating success (Coleman et al., 2007). Considering that Chinese Blackbirds copied human-made sounds and mating calls of Cuculidae birds (Fu et al., 2024), which is meaningless for them in function, we suggested that vocal mimicry in Chinese Blackbirds possibly supported the versatility hypothesis at least, which signals the quality of males (Dalziell et al., 2015). Chinese Blackbirds only mimicked the first phrase of the moped alarm because of two possible reasons. First, this phrase was easier to imitate than other sounds of mopeds (e.g., Fig. 3A–E). Second, this phrase was one of the most common sounds from mopeds in our study areas and thus was exposed to blackbirds more than other sounds.
We also found a geographic pattern of the alarms imitated by Chinese Blackbirds. Mimetic alarms from area 1 and area 2 were classified clearly by HCA and PCA (Figs. 1C and 5A-B): the mimetic alarms from area 1 possessed lower frequencies than those from area 2 according to Fig. 5C. One explanation for this geographic pattern is that Chinese Blackbirds may not only imitate the mimetic alarms from electric mopeds directly, namely the primary mimicry, but they also learn these sounds from close conspecific neighbors, namely the secondary mimicry (Fu et al., 2024; Gammon and Resh, 2024). Chinese Blackbirds can develop secondary mimicry (Gammon and Resh, 2024); for instance, they learn innovative mimicry of cuckoo calls from conspecific neighbors (Fu et al., 2024). One or a few males in area 1 might imitate an alarm phrase from electric mopeds directly, and then this mimetic sound was learned by the close conspecific neighbors. This might be the case for males from area 2 too. Males in area 2 imitated 1–2 notes of alarms or copied the locking/unlocking sounds, and hybridized these sounds with other song elements into one single song. Then this song type was shared by several neighbors. The initial imitators from different areas might copy different brands of mopeds (Fig. 5B, model alarms were also classified into two sets), and then the mimetic sounds were secondarily copied by other males. This process might lead to the geographical patterns of mimetic alarms. Additionally, if most of the males learned mimicry from conspecifics rather than electric mopeds, the inaccuracies of initial mimicry would be propagated.
Human-made sound mimicry indicates the adaptation of passerine mimics to urban environments. Rapid urbanization has driven the homogenization of avian community (Sun et al., 2022a; Deng et al., 2024), and reduced taxonomic diversity in urban areas (Ibáñez-Álamo et al., 2017), which probably reduces the available model species. Urban mimic species imitate human-made sounds to enrich their mimetic repertoires, though human-made sounds can be even more difficult to mimic than vocalizations from other bird species. In a similar case, male Albert's Lyrebirds mimicked fewer model species in small patches than those living in large patches, but they mimicked more vocalization types from each species to compensate for their mimetic repertoires partially (Backhouse et al., 2023). Additionally, Chinese Blackbirds also produced mimetic antitheft alarms of electric mopeds in different manners or arranged the mimetic units and species-specific song phrases in various temporal sequences. For instance, they produced mimetic alarms independently (Fig. 2D), incorporated the alarms with other mimetic vocalizations (Fig. 2E), or even arranged them with more complex sequences (Fig. 2F–Appendix Audio S1). Other mimics also combine or reconfigure song elements from various species into a new structure, such as Northern Mockingbirds (Gammon and Altizer, 2011). This may be a pathway to make a certain mimetic unit more complex in arrangement structures, which further facilitates the versatility of mimicry.
Our findings revealed that Chinese Blackbirds could mimic the sounds of electric mopeds, though the mimicry is not perfect by evaluation from a human perspective. Blackbirds imitated notes with less consistency, fewer notes per phrase, and lower frequencies than model alarms, indicating their mimicry was limited by cost or physical constraints. We also found that mimetic alarms from the two areas were different in acoustic structures, suggesting secondary mimicry might be general in Chinese Blackbirds. Human-made sound mimicry not only indicates the ability of oscine mimics to imitate challenging sounds, but also reflects the adaptation of urban birds to human-dominated environments. Considering vocal mimicry studies often focus on forest songbirds (Dalziell et al., 2015, 2021; Backhouse et al., 2022), we argue that further studies on urban species should be devoted. These studies should involve how many human-made sounds are imitated, the mimicry accuracy, and the correlation between urbanization progress and the proportion of human-made sounds in mimetic repertoires, which will broaden our insight into vocal mimicry from different environment types.
This study was conducted and approved by the Animal Welfare and Ethical Review Committee of Nanjing University (IACUC-2003001). No bird was captured or disturbed.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank Yuqi Sun for field works. We also would like to thank all owners of electric mopeds for their assistance and permission of the sound recordings.
Backhouse, F., Dalziell, A.H., Magrath, R.D., Welbergen, J.A., 2022. Higher-order sequences of vocal mimicry performed by male Albert's lyrebirds are socially transmitted and enhance acoustic contrast. Proc. R. Soc. B 289, 20212498.
Backhouse, F., Welbergen, J.A., Magrath, R.D., Dalziell, A.H., 2023. Depleted cultural richness of an avian vocal mimic in fragmented habitat. Divers. Distrib. 29, 109–122.
Benedict, L., Charles, A., Brockington, A., Dahlin, C.R., 2022. A survey of vocal mimicry in companion parrots. Sci. Rep. 12, 20271.
Budaev, S.V., 2010. Using principal components and factor analysis in animal behaviour research: caveats and guidelines. Ethology 116, 472–480.
Coleman, S.W., Patricelli, G.L., Coyle, B., Siani, J., Borgia, G., 2007. Female preferences drive the evolution of mimetic accuracy in male sexual displays. Biol. Lett. 3, 463–466.
Cortopassi, K.A., Bradbury, J.W., 2000. The comparison of harmonically rich sounds using spectrographic cross-correlation and principal coordinates analysis. Bioacoustics 11, 89–127.
Cramer, E.R.A., 2012. Vocal deviation and trill consistency do not affect male response to playback in house wrens. Behav. Ecol. 24, 412–420.
Dalziell, A.H., Magrath, R.D., 2012. Fooling the experts: accurate vocal mimicry in the song of the superb lyrebird, Menura novaehollandiae. Anim. Behav. 83, 1401–1410.
Dalziell, A.H., Maisey, A.C., Magrath, R.D., Welbergen, J.A., 2021. Male lyrebirds create a complex acoustic illusion of a mobbing flock during courtship and copulation. Curr. Biol. 31, 1970–1976.e1974.
Dalziell, A.H., Welbergen, J.A., 2016. Mimicry for all modalities. Ecol. Lett. 19, 609–619.
de Kort, S.R., Eldermire, E.R.B., Valderrama, S., Botero, C.A., Vehrencamp, S.L., 2009. Trill consistency is an age-related assessment signal in banded wrens. Proc. R. Soc. B 276, 2315–2321.
del Hoyo, J., Collar, N., Christie, D.A., 2020. Birds of the World. Version 1.0. Cornell Lab
of Ornithology, Ithaca, NY, USA.
Deng, J., Zhu, Y., Luo, Y., Zhong, Y., Tu, J., Yu, J., et al., 2024. Urbanization drives biotic homogenization of the avian community in China. Integr. Zool. 00, 1–13.
Fu, C., Wang, X., Backhouse, F., Li, Z., 2024. Learning mimetic cuckoo call innovations from neighbors in a Chinese songbird. Sci. Rep. 14, 24079.
Gammon, D.E., Altizer, C.E., 2011. Northern Mockingbirds produce syntactical patterns of vocal mimicry that reflect taxonomy of imitated species. J. Field Ornithol. 82, 158–164.
Gammon, D.E., Corsiglia, A.M., 2019. Mockingbirds imitate frogs and toads across North America. Behav. Process. 169, 103982.
Gammon, D.E., Resh, G.E., 2024. Do vocal mimics learn their mimetic songs from heterospecifics or conspecifics? Behaviour 161, 441–475.
Hausberger, M., Jenkins, P.F., Keene, J., 1991. Species-specificity and mimicry in bird song: are they paradoxes? A reevaluation of song mimicry in the European starling. Behaviour 117, 53–81.
Ibáñez-Álamo, J.D., Rubio, E., Benedetti, Y., Morelli, F., 2017. Global loss of avian evolutionary uniqueness in urban areas. Glob. Change Biol. 23, 2990–2998.
Jůzlová, Z., Riegert, J., 2012. Vocal mimicry in the song of the icterine warbler, Hippolais icterina (Sylviidae, Passeriformes). Folia Zool. 61, 17–24, 18.
K. Lisa Yang Center for Conservation Bioacoustics, 2024. Revan Pro: Interactive Sound
Analysis Software. [Computer software] Version 1.6.5. The Cornell Lab of
Ornithology, Ithaca, NY. .
Kelley, L.A., Healy, S.D., 2011. The mimetic repertoire of the spotted bowerbird Ptilonorhynchus maculatus. Naturwissenschaften 98, 501–507.
Lambrechts, M., Dhondt, A.A., 1988. The anti-exhaustion hypothesis: a new hypothesis to explain song performance and song switching in the great tit. Anim. Behav. 36, 327–334.
Liu, X., Li, Z., 2024. Urban bird nest building on man-made structures: a review. Zool. Res. Divers. Conserv. 1, 273–281.
National Bureau of Statistics, 2023. Chinese Statistical Yearbook. Chinese Statistical Press, Beijing.
Paradis, E., Schliep, K., 2019. Ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528.
Pavlacky, D.C., Possingham, H.P., Goldizen, A.W., 2015. Integrating life history traits and forest structure to evaluate the vulnerability of rainforest birds along gradients of deforestation and fragmentation in eastern Australia. Biol. Conserv. 188, 89–99.
Phillips, J.N., Derryberry, E.P., 2017a. Equivalent effects of bandwidth and trill rate: support for a performance constraint as a competitive signal. Anim. Behav. 132, 209–215.
Phillips, J.N., Derryberry, E.P., 2017b. Vocal performance is a salient signal for male-male competition in White-crowned Sparrows. Auk 134, 564–574.
R Core Team, 2023. R: a Language and Environment for Statistical Computing. R
Foundation for Statistical Computing. .
Revelle, W., 2024. Psych: procedures for psychological, psychometric, and personality research. R package version 2.4.6.26. .
Sakata, J.T., Vehrencamp, S.L., 2012. Integrating perspectives on vocal performance and consistency. J. Exp. Biol. 215, 201–209.
Sierro, J., de Kort, S.R., Hartley, I.R., 2023. Sexual selection for both diversity and repetition in birdsong. Nat. Commun. 14, 3600.
Sierro, J., de Kort, S.R., Riebel, K., Hartley, I.R., 2022. Female blue tits sing frequently: a sex comparison of occurrence, context, and structure of song. Behav. Ecol. 33, 912–925.
Sprau, P., Roth, T., Amrhein, V., Naguib, M., 2013. The predictive value of trill performance in a large repertoire songbird, the nightingale Luscinia megarhynchos. J. Avian Biol. 44, 567–574.
Sun, B., Lu, Y., Yang, Y., Yu, M., Yuan, J., Yu, R., et al., 2022a. Urbanization affects spatial variation and species similarity of bird diversity distribution. Sci. Adv. 8, eade3061.
Sun, C., Zhang, C., Lucas, J.R., Gu, H., Feng, J., Jiang, T., 2022b. Vocal performance reflects individual quality in male Great Himalayan leaf-nosed bats (Hipposideros armiger). Integr. Zool. 17, 731–740.
Sun, S., Wang, P., 2023. The association between e-moped usage and residents' subjective well-being: a case study of Shanghai, China by using Bayesian network. Transport. Plann. Technol. 46, 976–997.
ten Cate, C., Fullagar, P.J., 2021. Vocal imitations and production learning by Australian musk ducks (Biziura lobata). Philos. Trans. R. Soc. B-Biol. Sci. 376, 20200243.
Wang, L., Lam, T.T., Xu, S., Dai, Z., Zhou, L., Feng, T., et al., 2020. Treeio: an R package for phylogenetic tree input and output with richly annotated and associated data. Mol. Biol. Evol. 37, 599–603.
Yu, G., Smith, D.K., Zhu, H., Guan, Y., Lam, T.T., 2017. GGTREE: an R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 8, 28–36.
Zann, R., Dunstan, E., 2008. Mimetic song in superb lyrebirds: species mimicked and mimetic accuracy in different populations and age classes. Anim. Behav. 76, 1043–1054.
Zhao, Z., Borzée, A., Li, J., Chen, S., Shi, H., Zhang, Y., 2023. Urban bird community assembly mechanisms and driving factors in university campuses in Nanjing, China. Animals 13, 673.
Zheng, G.M., 2023. A Checklist on the Classification and Distribution of the Birds of China, fourth ed. Science Press, Beijing.
Zollinger, S.A., Suthers, R.A., 2004. Motor mechanisms of a vocal mimic: implications for birdsong production. Proc. R. Soc. B 271, 483–491.
Table
1.
The factor loadings of principal component analysis for nine acoustic parameters of model and mimetic alarms. Loadings >0.40 are shown in bold.
Table
2.
Mean (SD) and differences between model and mimetic alarms for each acoustic parameter. N indicates the number of individuals; np indicates the number of phrases; nn indicates the number of notes.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds