
Figure
1.
The Xinjiang Ground Jay in the Lopnur Desert (Photo by Ming Ma)
| Citation: |
Xinsen Wei, Zihui Zhang. 2023: Model-based assessment of muscle forces and strain distribution in the femur of Cabot’s Tragopans (Tragopan caboti). Avian Research, 14(1): 100149. DOI: 10.1016/j.avrs.2023.100149

|
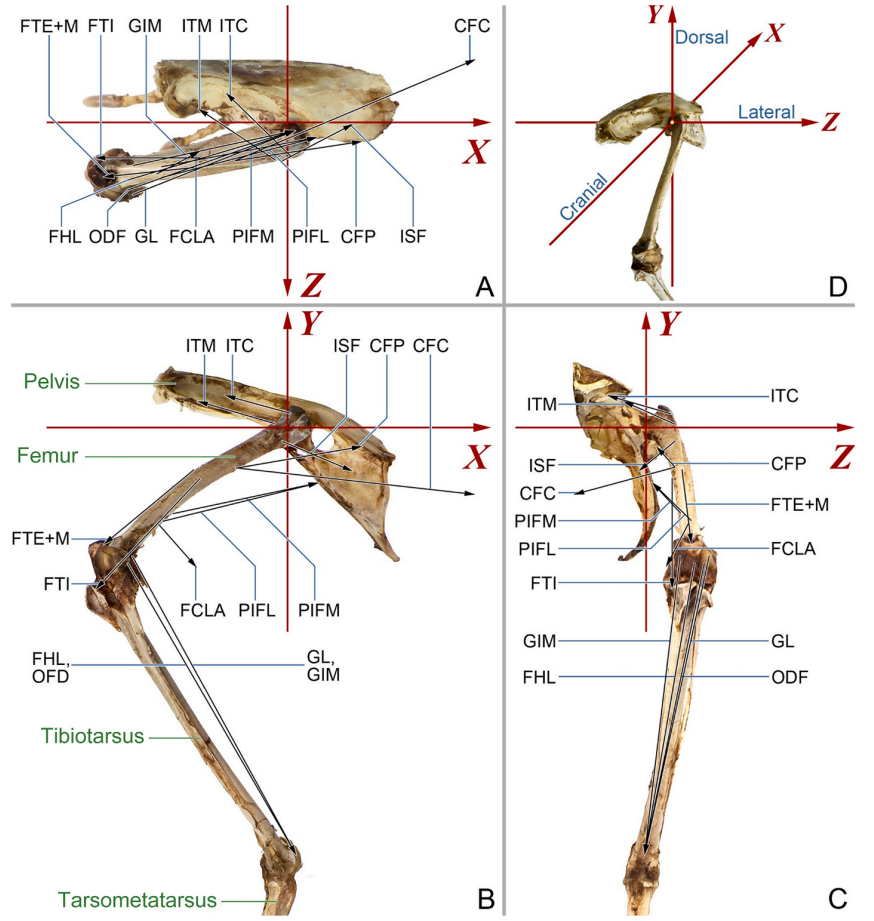
The hindlimbs play a crucial role in bird locomotion, making the biomechanical properties of the musculoskeletal system in these limbs a focal point for researchers studying avian behaviour. However, a comprehensive analysis of the mechanical performance within the long bones of hindlimbs during locomotion remains lacking. In the present study, the strain and deformation of the femur of Cabot’s Tragopans (Tragopan caboti) were estimated. We employed inverse simulation to calculate the force and moment of femoral muscles during mid-stance terrestrial locomotion and conducted finite element analysis to calculate femoral strain. Results showed that during mid-stance, the femur experiences combined deformation primarily characterized by torsion, bending, and compression. It emphasises the importance of considering the influence of varying loads on bone adaptation when investigating bone form-function relationships. Muscles were found to play a significant role in offsetting joint loads on the femur, subsequently reducing the deformation and overall strain on the bone. This reduction enhances femoral safety during locomotion, allowing birds to meet mechanical demands while maintaining a lightweight bone structure. Notably, the M. iliotrochantericus caudalis significantly reduces torsional deformation of the proximal femur, protecting the vulnerable femoral neck from high fracture risk induced by rotation load. Given that the femur torsion during terrestrial locomotion in birds is associated with changes in hindlimb posture due to their adaptation to flight, the characteristics of M. iliotrochantericus caudalis may provide insight into the locomotor evolution of theropods and the origin of avian flight.
Among the four species of ground jays (Podoces) in the world, two are found in the west of China (Qian et al., 1965; Cheng, 1987): the Xinjiang Ground Jay (P. biddulphi) (Fig. 1) and the Mongolian Ground Jay (P. hendersoni). Xinjiang Ground Jays occur only in the Taklimakan Desert, the southern part of Xinjiang. Since the species was established by A. Hume in 1874, little has been known of its status and ecology. The current essay describes such information based on a long-term field survey.
The ground jays are residents at the center of the Taklimakan Desert (37–42°N, 77–94°E, 790–1500 m elevations; Fig. 2). Most of distribution range of the species falls within Xinjiang, with a few extending to the east, e.g. the Qaidam Basin in Qinghai Province and Dunhuang in Gansu Province (Collar et al., 2001; Sun and Li, 2009). Interestingly, Mongolian Ground Jays are distributed around the range of Xinjiang Ground Jays (Fig. 2). Such a pattern should be a result of inter-species competition. There is evidence showing that the ground jays are expanding their range from west to east during recent decades (Ma, 2010).

Ground jays are well adapted to desert and semi-desert regions (Ludlow and Kinnear, 1933). Compared with crows or choughs, ground jays are sandy in plumage, presumably providing protection from desert predators (Londei, 2004). As their names indicate, ground jays spend much of their time on the ground. The strong legs should be adaptive to the habit. However, ground jays nest in shrubs and trees, a characteristic similar to that followed by the Corvids species.
Information on breeding ecology of Xinjiang Ground Jays is very limited. A total of 20 ground jay nests were recorded in Niya and Qarqan from 2003 to 2004. The birds placed their nests on the small desert-poplar tree Populus diversifolia and Tamarix spp. bushes, averaging 1.09 ± 0.15 m (range = 0–2.30 m, n = 18) above the ground (Fig. 3). The nests were composed of sheep wool, camel's wool, horse's hair, dead leaves, dry grass, and the soft cottony growth of reeds, with poplar skin, twigs and small sticks being lined at the base. The external diameter of the nest is 35.75 ± 2.30 cm (range = 16–55 cm, n = 16), the internal diameter of the nest is 12.82 ± 0.85 cm (range = 9–20 cm, n = 14), and depth of the nest is 9.50 ± 1.00 cm (range = 5–16 cm, n = 13) and the height is 20.88 ± 1.33 cm (range = 12–35 cm, n = 16) (Fig. 4a). Clutch size varied between 1 and 3 eggs (1.89 ± 0.31 eggs, n = 9), the diameters of the eggs are 32.88 ± 0.83 mm × 23.48 ± 0.09 mm, and the weight of egg is 8.33 ± 0.88 g (n = 4). The color of egg is pale green and grayish white with brown spots scattered all over the surface, rather more densely at the broad end (Fig. 4b). The parents fed the young (Fig. 4c) more than 42 times during one day for one nest located in the middle March (Ma, 2004). After fledging, family flocks of 4 to 6 birds were encountered between early May and July. These data suggested that the ground jays laid eggs from late February to April.
Based on the transect counting conducted from 1988 to 2011, the density of ground jays was to be 3–5 pairs per 100 km2. It was estimated to have 4100–6700 pairs of ground jays over the species' range of 135000 km2.
The ground jay populations seemed to drop during recent decades (Grimmett, 1991; Madge and Burn, 1994; Ma, 1998). However, with the intensified desertification in western China, the jays have a tendency to expand to the east.For example, there were some of new records in Gansu and Qinghai provinces (Collar et al., 2001; Sun and Li, 2009; Ma, 2010). Arguably, this bird is an indicator species of desertification and climate change.
Human activities such as oil industry, ecotourism, land exploitation and overgrazing should be responsible for the population decline (Ma, 2001; Ma and Kwok, 2004). The restricted range and special requirements for nesting habitats suggest an urgent conservation need for this species. Now, although the bird is classified as "near-threatened" (Collar et al., 2001), no protection measures have been in practice (Zheng and Wang, 1998).
The research is supported by the Science Supporting Project of National Ministry of Science and Technology (2008BAC39B04) and the National Natural Science Foundation of China (30270211, 30470262, 30970340). Sincere thanks are due to all participants in the field work, especially to Jinghe Gu, Kwok Hon Kai, Zexin Jia, Batuerhan, Eugene Potapov, Chuanbo Wang, Feng Xu, Yiqun Wu, Mike Kilburn, Geoff Carey, Richard Lewthwaite, Sebastien Lepetz and Paul Leader. Gratitudes are also given to Prof. Xin Lu from Wuhan University who helps revise the earlier manuscript very carefully.
|
Currey, J.D., 2002. Bones: Structure and Mechanics. Princeton University Press, Princeton.
|
|
Goodno, B.J., Gere, J.M., 2018. Mechanics of Materials, ninth ed. Cengage Learning, Boston.
|
|
Khassetarash, A., Haider, I., Baggaley, M., Edwards, W.B., 2022. Tibial strains during prolonged downhill running: a finite element analysis. J. Biomech. Eng. 145, 041007.
|
|
Mendez, J., Keys, A., 1960. Density and composition of mammalian muscle. Metabolism 9, 184–188.
|
|
Young, V.K.H., Wienands, C.E., Wilburn, B.P., Blob, R.W., 2017. Humeral loads during swimming and walking in turtles: implications for morphological change during aquatic reinvasions. J. Exp. Biol. 220, 3873–3877.
|
|
Zhang, Z., Zheng, G., 1996. A study on the appendicular myology of the Cabot’s tragopan. In: Society, C.O., et al. (Ed.), Studies of Chinese Ornithology. China Forestry Publishing House, Beijing, pp. 210–216. (in Chinese with English abstract).
|
Figures(5) / Tables(1)





 DownLoad:
DownLoad:


 Email Alerts
Email Alerts RSS Feeds
RSS Feeds