Santi Guallar, Javier Quesada. 2023: Moult intensity constraints along the complete moult sequence of the House Sparrow (Passer domesticus). Avian Research, 14(1): 100125. DOI: 10.1016/j.avrs.2023.100125
Citation:
Santi Guallar, Javier Quesada. 2023: Moult intensity constraints along the complete moult sequence of the House Sparrow (Passer domesticus). Avian Research, 14(1): 100125. DOI: 10.1016/j.avrs.2023.100125
Santi Guallar, Javier Quesada. 2023: Moult intensity constraints along the complete moult sequence of the House Sparrow (Passer domesticus). Avian Research, 14(1): 100125. DOI: 10.1016/j.avrs.2023.100125
Citation:
Santi Guallar, Javier Quesada. 2023: Moult intensity constraints along the complete moult sequence of the House Sparrow (Passer domesticus). Avian Research, 14(1): 100125. DOI: 10.1016/j.avrs.2023.100125
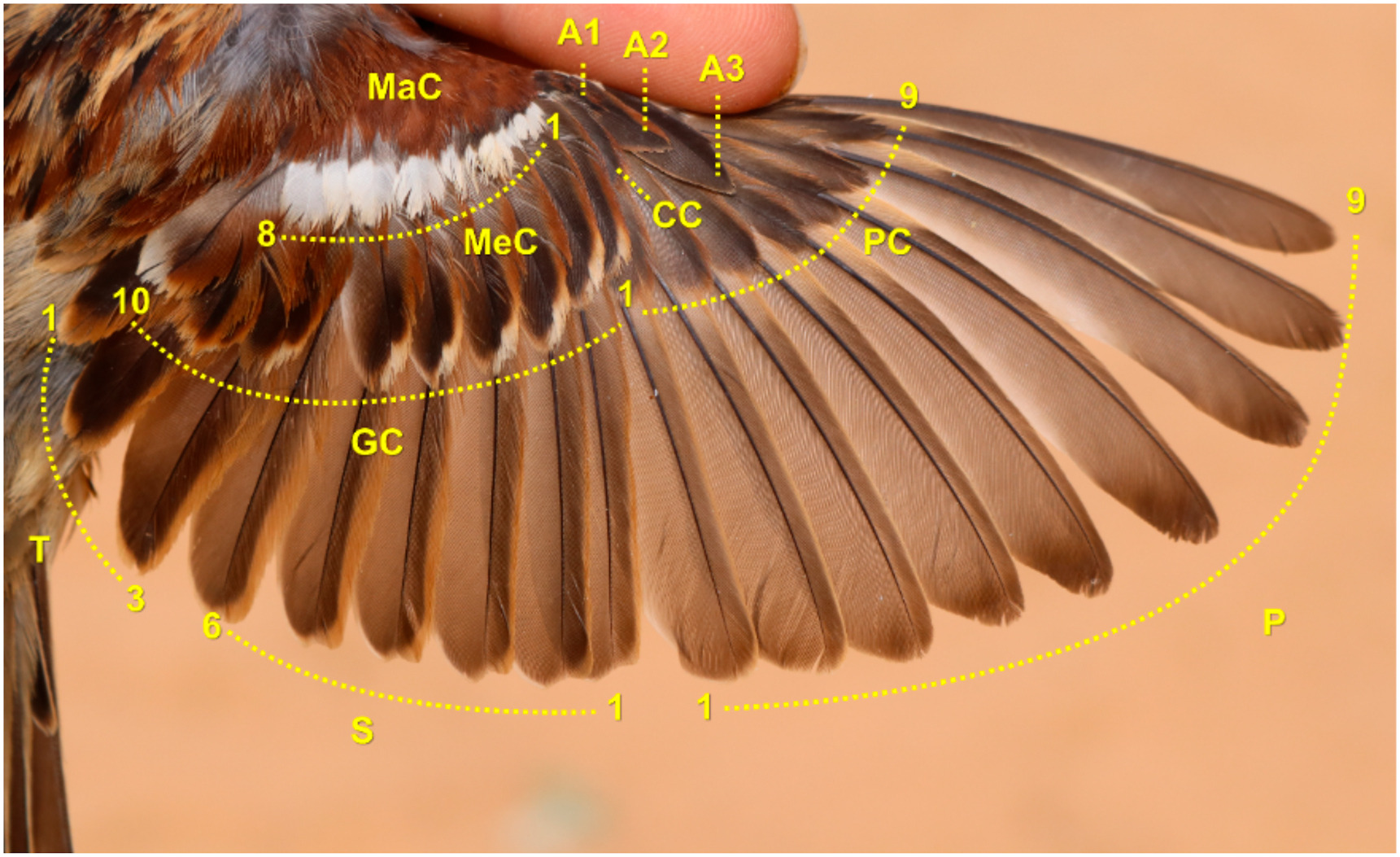
Sequence and intensity are two essential components of bird moult. While the moult sequences of remex tracts are highly homogenous across passerines, other tracts apparently show a high variability. Moreover, order of moult activation among tracts are insufficiently known. Likewise, dynamics of moult intensity as moult progresses remains poorly known. Here, we provide detailed quantitative description of moult sequence and intensity in the House Sparrow (Passer domesticus). To understand their role, we tested two hypotheses on the: 1) protection function of moult sequence, and 2) aerodynamic and physiological constraints on moult intensity. We scored percentage growth of 313 captured sparrows using the mass of the feathers of each tract (also length for remiges) to monitor moult intensity throughout the complete moult progress, which is defined as the fraction of new and growing feathers in a moulting bird relative to the total plumage. Moult sequence was highly variable both within wing coverts and among feather tracts, with moult sequence differing among all birds to some degree. We only found support for the protection function between greater coverts and both tertials and secondaries. Remex-moult intensity conformed to theoretical predictions, therefore lending support to the aerodynamic-constraint hypothesis. Furthermore, remex-moult speed plateaued during the central stages of moult progress. However, overall plumage-moult speed did not fit predictions of the physiological-constraint hypothesis, showing that the remex moult is only constrained by aerodynamics. Our results indicate that aerodynamic loss is not simply the inevitable effect of moult, but that moult is finely regulated to reduce aerodynamic loss. We propose that the moult of the House Sparrow is controlled through sequence and intensity adjustments in order to: 1) avoid body and wing growth peaks; 2) fulfil the protection function between some key feather tracts; 3) reduce detrimental effects on flight ability; 4) keep remex sequence fixed; and 5) relax remex replacement to last the whole moult duration.
Most signals are thought to be reliable based on the underlying logic that receivers only respond to signals that are sufficiently reliable (Maynard and Harper 2003; Searcy and Nowicki 2005). If a signal is always unreliable, the receivers would ignore that signal. Thus, the signaling systems would not exist at all. When the signaler's and receiver's interests are opposed (e.g., in male-male competition for resources), there are two main explanations for the reliability of signals. First, some signals are reliable because those signals are forced to be honest due to physiological or anatomical constraints on signal production (Maynard and Parker 1976). For example, the dominant frequencies of calls used in male-male conflict in some frogs belongs in this category; the frequency is determined by the size of the vocal apparatus, which is determined by body size, and the dominant frequency is thus constrained to reflect reliable information about body size (Martin 1971, 1972). A performance constraint on the trilled vocalizations in Emberizidae is an additional example: maximal values of frequency bandwidth and trill rates are limited by motor constraint (Podos 1996, 1997). The second explanation for signal reliability is based on costs. Costs that stem from signaling can prevent deceit if the signaler's fitness is decreased due to cheating (Grafen 1990). Birds often use vocal signals in territorial disputes, and the vocal signal used here might be associated with escalated contests (Searcy and Beecher 2009). If a weak individual cannot bear an escalated contest, reliable vocal signals could guarantee the cost of a conceivable escalated contest (Vehrencamp 2000; Molles and Vehrencamp 2001).
Although signals are often honest, unreliable signaling can exist in signaling systems (Maynard and Harper 2003; Searcy and Nowicki 2005). Unreliable signaling can be defined by departures from the typical correlation between the signal and an attribute of the signaler (Hauser 1996). For example, If signal a is typically correlated with action A, unreliable signaling occurs when an individual signals a but does not perform A (i.e., over-signaling) or does not signal a but does perform A (i.e., under-signaling). Depending on the value of the contested resource and the expected cost of an escalated contest for the signaler, models predict that unreliable signaling can be a useful strategy for some individuals or in some context (Szamado 2000; Szalai and Szamado 2009). For example, although song type matching (i.e., singing to the rival with the same song that the rival has just sung) has long be seen as an aggressive signal in song sparrows during male-male conflict (Stoddard et al. 1992), this signal can reflect the signaler's expectation of an escalated contest in the initial of territory conflict but fail to predict direct attack (Searcy et al. 2006; Akcay et al. 2013). Because the reliability of a signal is context dependent, testing the signal in different contexts might aid the understanding of both the function of the signal and the evolution of the signaling system.
Soft song is characterized by markedly lower amplitudes than normal song (here termed broadcast song) and has long been observed in many birds (Dabelsteen et al. 1998; Morton 2000). Recent studies have confirmed that soft song is an aggressive signal; soft song can predict aggressive escalation in both passerine (Searcy et al. 2006; Ballentine et al. 2008; Hof and Hazlett 2010; Akcay et al. 2011) and non-passerine birds (Rek and Osiejuk 2011). Additionally, playback experiments with mounted specimens have shown that soft song is a reliable aggressive signal in male-male conflict (Searcy and Beecher 2009; van Staaden et al. 2011; Searcy et al. 2014). In a previous study, we found that the relationship between soft song production and subsequent attack is significant in the Brownish-flanked Bush Warbler Cettia fortipes (Xia et al. 2013b). Here, we examined the correlation between soft song production and subsequent attack in different male-male conflict contexts in this species.
The brownish-flanked bush warbler is a small, furtive passerine found throughout Southeast Asia. During the breeding season, the males sing clear, high-pitched broadcast songs from dense undergrowth (del Hoyo et al. 2006; Kennerley and Pearson 2010). The males have a small repertoire of broadcast songs (typically two or three songs) and always sing the broadcast song types in alternating patterns (e.g., a-b-a-b; Xia et al. 2013a). Compared to broadcast song, soft song in this species is characterized by lower amplitude, a significantly lower minimum frequency, more notes, a longer duration, and a higher note rate (Xia et al. 2013b). Thus, soft song is easily distinguished from broadcast song in the field. Based on our observations, broadcast song is always used in spontaneous singing and after expelling an intruder, while soft song is used during encounters between rival males. In a previous playback study (Xia et al. 2013b), we used mounted specimens positioned within the territory of subject males and conducted playback trails of broadcast songs and found that six of 25 territorial owners attacked the mounted specimen, and all attacking subjects generated soft songs immediately prior to the attacks. However, the context of the playback experiment of Xia et al. (2013b) is uncommon in the field because in contrast to broadcast song, soft song was always produced by the intruder when in close contact with the territory owner. The first aim of this study was to investigate whether soft song and subsequent attack are correlated in the territorial owner when the intruder produces soft song. The context of this study is more similar to that of the field when the territory owner closely encounters the intruder. Thus, this context can provide additional information about the function of soft song for birds.
The second aim of this study was to investigate the cost of soft song in the Brownish-flanked Bush Warbler. For signals with no obvious physiological or anatomical constraints, cost should be the most probable explanation for the stability of the signaling system (Vehrencamp 2000; Molles and Vehrencamp 2001). A previous study showed that soft song can induce male brownish-flanked bush warbler to spend significantly more time near the loudspeaker (Xia et al. 2013b). We hypothesized that closeness is a cost to the signaler in the experiment detailed by Xia et al. (2013b) based on the underlying logic that increasing closeness increases the possibility of attack. However, this underlying logic has not been verified in this species. In the present study, we show direct evidence of cost to the soft song signaler; i.e., soft songs induced more attacks.
Methods
Study area and species
During May 2013 we conducted experiments on a population of brownish-flanked bush warblers in the Dongzhai National Nature Reserve in the Henan province of southern China (31.9°N, 114.3°E). The males in this population have been monitored, color-banded and recorded as a part of a long-term study since 2007. Brownish-flanked bush warblers defend territories from March to July. Most of the territories are characterized by dense bushes that are dominated by Camellia sinensis tea plants; thus, the territory boundaries are typically well defined by the habitat.
Song stimuli
The songs used in the playback experiments were recorded using a TASCAM HD-P2 portable digital recorder (Tascam Co., Japan) and a Sennheiser MKH416P48 external directional microphone (Sennheiser Co., Germany) at a sampling rate of 44.1 kHz and a sample size of 16 bits. For the playback trials, we randomly selected five broadcast songs that were recorded from five different males during the 2011 breeding season and five soft songs that were randomly selected from five different males recorded during the 2012 breeding season. We used Goldwave 5.25 (GoldWave Inc., Canada) to adjust the broadcast song rate to one song every 10 s, which is similar to the song rate of this species. For the soft songs, we repeated the recordings to achieve a total time of 3 min because the majority of the soft song recordings were shorter than 3 min. Using a CEL-240 sound level meter (Casella Co., USA), the speaker volume for the playback trials of the broadcast and soft songs were adjusted to 80 dB and 60 dB, respectively, at a distance of 1 m above the speaker. This amplitude approximates the natural amplitude of song in the field as evaluated by ear by observers. In total, we used ten unique 3-min playback stimuli that included five broadcast songs and five soft songs. Similar playback stimuli have been used in a previous playback study of the species (see Xia et al. 2013b).
Playback procedure
We used mounted specimens of four male brownish-flanked bush warblers obtained with permission from the Museum of Beijing Normal University, and we randomly selected one for each subject male. Prior to each playback trial, we located each subject male by its spontaneous singing behavior. We positioned a single mounted specimen on the ground concealed under bushes within the territory of each subject male at approximately 10 m from the singing spot. The specimens were always placed away from the territorial boundary to decrease the effects on neighbors. Playback was conducted using uncompressed.wav files stored on a TeclastX18Mp3 player (Teclast Co., Shenzhen, China) connected to a Senway loudspeaker (Senway Amplifier Co., Shenzhen, China). The loudspeaker was placed next to the mounted specimen and concealed with grass or dead leaves.
There were 51 subject males used in this study. Each subject male was exposed to one playback trial. The type of playback (either broadcast or soft song playback) was determined by a coin toss for the first subject and was then alternated between broadcast song and soft song for the subsequent subject males. The specific broadcast or soft song playback stimuli were randomly selected for each subject male. Most of the males in this population have been color-banded, which allowed us to ensure that the playback stimuli were not recorded from the subject male or its immediate neighbors. Two observers, positioned approximately 10 m away from the mounted specimen, recorded the behavioral responses of the subject males. We stopped playback if the subject male began to physically attack the mounted specimen. Otherwise, the experiment continued for the 3 min duration. If a subject male did not appear within 3 m during the 3 min playback, we abandoned the trial because we could not be sure that the subject male had seen the mounted specimen. If the 3 m criterion was met, we recorded the attack latency, which was defined as the time from which the male first appeared within 3 m of the mounted specimen to attack (defined as any physical contact with the mount specimens). Due to difficulties in observing some behaviors (e.g., wing quivering) in the dense vegetation of the birds' habitat, we only recorded whether soft song was produced for each subject male. Because most attack latencies were < 10 s in our previous study (Xia et al. 2013b), we were only concerned with signals that occurred within 10 s prior to the attack. For those subjects that did not attack, we focused on the signals in the last 10 s within 3 m of the mounted specimens during 3 min playback. We obtained data from 51 subjects that included 26 subjects in the broadcast songs playback group and 25 subjects in the soft songs playback group.
Data analysis
We used Fisher's exact test to compare the numbers of attacks by the subject males between the broadcast song playback group and the soft song playback group. After we removed the data for the subject males that did not attack the mount specimen, a t test was used to compare the attack latency between the broadcast song playback group and soft song playback group. To test the relationship between soft song and subsequent attack, we used Fisher's exact tests for the broadcast song playback group and the soft song playback group, respectively.
All data are expressed as the mean ± the SE, and differences were considered significant at P < 0.05. The data were analyzed using SPSS v.20 (IBM Co., New York, NY, USA). Permission for this study was granted by the National Bird-banding Center of China (license number H20110042) and the Dongzai National Nature Reserve (license number 2011002).
Results
The relationship between soft song and subsequent attack was significant in the broadcast song playback group (Fisher exact test, P = 0.046) but not in the soft song playback group (Fisher exact test, P = 0.202; Table 1). Both under-signaling and over-signaling subject males were observed. In broadcast song playback group, 15.4% subject males attacked without producing soft song (under-signaling) and 11.5% subject males produced soft song but did not attack (over-signaling); in soft song playback group, 28.0% subject males attacked without producing soft song (under-signaling) and 8.0% subject males produced soft song but did not attack (over-signaling) (Table 1).
Table
1.
Number of subject males to 3 min of playback of broadcast or soft songs
The soft song playback group evoked significantly more attacks than did the broadcast song playback group (Fisher exact test, P = 0.025; Table 1), and the attack latency was significantly shorter in the soft song playback group (23.2 ± 7.9 s) than in the broadcast song playback group (59.9 ± 16.7 s; t test, t26 = 2.252, P = 0.033; Figure 1).
Figure
1.
Attack latency (mean ± SE) of the subject males in the playback experiment.
Attack latency was defined as the time between the first appearance of the subject male within 3 m of the mounted specimens and the time of physical contact with the mount specimen. The attack latency was significantly shorter in soft songs playback group than in the broadcast song playback group (t test, t26 = 2.252, P = 0.033).
Although the signal must be sufficiently reliable to maintain a stable signaling system, there is room for cheating as a strategy for some individuals or in some context (Szamado 2000; Szalai and Szamado 2009). In this study, we found that the correlation of the subject males' soft songs and subsequent attacks was significant in the broadcast song playback group but not in the soft song playback group. The reliability of an aggressive signal requires a correlation between the signal and an escalation toward physical fighting on the part of the signaler (Hauser 1996; Searcy and Nowicki 2005). Our data show that there was less reliability in the soft song playback group.
Can the subject males benefit from the unreliable signaling? There were two males in the broadcast song playback group and three males in the soft song playback group that produced soft song but did not attack the intruder (i.e., the mounted specimen). The potential benefit of this over-signaling is intimidation of the intruder that could cause the intruder to concede (Searcy and Nowicki 2005; Maynard and Harper 2003). Comparison of the attack rates of the soft song playback group (72.0%) and the broadcast song playback group (38.5%) indicates that soft songs from the intruder (mounted specimen) did not cause more territory owners to concede. However, we should note the asymmetry between the intruder and the territory owner (Maynard and Price 1973). Territory is always more valuable for the territory owner, thus it is not easy for the territory owner to be intimidated by the soft song of an intruder. Whether over-signaling benefits the territory owner remains an open question because the intruder (mounted specimen) could not retreat in our experiment. There were four males in the broadcast song playback group and seven males in soft song playback group that did attack the intruder (mounted specimen) but did not produce soft song. We believe that the potential benefit acquired by these under-signaling subject males was gaining the initiative in fighting. Based on this hypothesis, we infer that under-signaling might occur more often when a fight is inevitable. Our data support this inference; more under-signaling subject males were observed in the soft song playback group (28.0%) than in the broadcast song playback group (15.4%). In the future, we will demonstrate that initiating attack is beneficial in fights, and we will then fully understand the benefit acquired by the under-signaling subject males.
Limited reliability for soft song is also found in other species, i.e. attack was not correctly predicted in about one third of the Song Sparrow Melospiza melodia males (Akcay et al. 2013). Similarly, under-signaling is more common than over-signaling (Searcy et al. 2006; Akcay et al. 2013). As how responses to intrusion may rely on attributes of responding animals themselves (Moseley et al. 2013), high quality individuals may adopt under-signaling strategy without bothering to waste time in signaling (Searcy et al. 2013). To test this hypothesis, fighting ability or/and aggressive intentions will be compared between under-signaling males and other males in future work.
The cost of soft song
Cost is crucial for the maintenance of aggressive signaling systems (Vehrencamp 2000; Molles and Vehrencamp 2001). If there is no cost for aggressive signaling, it is advantageous for individuals produce the maximal levels such signals during conflicts. If all individuals signal maximally, it will be difficult for the receivers to judge the signaler's aggressive intentions based on the information contained in that signal. Thus, the receivers would ignore the signal, and the signaling system could not exist.
Among several potential cost concern soft song (summarized in Akcay et al. 2011; Akcay and Beecher 2012), receiver-retaliation cost get most support (Ballentine et al. 2008; Hof and Hazlett 2010; Anderson et al. 2012; Templeton et al. 2012). Receiver-retaliation cost posits that soft song increases the likelihood of the receiver retaliating aggressively. In a previous study, we found that soft song induces subject males to spend more time near the sound source (i.e., loudspeaker; Xia et al. 2013b) and inferred that being closer might indicate an increased possibility of attack. Recent studies have found that some behaviors that were previously thought to reflect attack intent are not actually correlated with attack (Ballentine et al. 2008; Hof and Hazlett 2010; Searcy et al. 2006). Thus, it is insufficient to assume that the closeness of the receiver is a cost of to the soft song signaler (Hof and Hazlett 2010; Hof and Podos 2013). The present study showed that when soft song was played back, more subject males conducted attacks of the mounted specimen with shorter attack latencies. The greater number of subject males attacking on mounted specimen indicates greater harm due to fighting to the signaler, and the shorter attack latency indicates that the signaler had less time for retreat. Both these factors provide direct evidence regarding receiver-retaliation cost of soft song; i.e., signalers that produce soft song put themselves at risk for fighting.
Besides receiver-retaliation cost (Anderson et al. 2012; Templeton et al. 2012; Searcy et al. 2014), there is cost from potential intruder in the song sparrow: as soft songs are low in amplitude, a potential intruder might fail to hear a male who uses soft song in conflict and thus be more likely to intrude on the soft song signaler's territory (Searcy and Nowicki 2006). As increasing evidence indicates that territorial conflicts can be eavesdropped on by other potential rivals (Akcay et al. 2010; Sprau et al. 2012), future work should test whether brownish-flanked bush warbler soft song signalers experience a greater number of intrusions.
Conclusions
In this work, we showed the receiver-dependent costs entailed by the soft song of the Brownish-flanked Bush Warbler; i.e., the signaler that produces soft song suffer more and quicker attacks from the receiver. We observed both over-signaling and under-signaling male Brownish-flanked Bush Warblers. In simulated intrusion that coupled playback of soft song with a mount specimen, a greater number of territory owner directly attacked without producing soft song (under-signaling). We suggest that such under-signaling males might benefit from taking the initiative in fights.
Competing interests
The authors declare that they have no competing interests.
Acknowledgements
This study was supported by National Natural Science Foundation of China (No.31172098) and the China Postdoctoral Science Foundation (No.2014 M550026). We would like to thank Jiayu LIU for her help on field work.
Authors' contributions
YYZ and CWX conceived and designed the experiments. CWX, DPW, BYL performed the experiments. CWX analyzed the data. YYZ, CWX, HL wrote or revise the manuscript. All authors read and approved the final manuscript.
Bååth, R., 2016. Package 'bayesboot'. An Implementation of Rubin's (1981) Bayesian Bootstrap. Version 0.2.1. .
Bensch, S., Grahn, M., 1993. A new method for estimating individual speed of molt. Condor 95, 305–315.
Bridge, E.S., 2003. Effects of simulated primary moult on pigeon flight. Ornis Fenn. 80, 121–129.
Bridge, E.S., 2008. How does imping affect wing performance? J. Wildl. Rehabil. 29, 4–9.
Chai, P., 1997. Hummingbird hovering energetics during moult of primary flight feathers. J. Exp. Biol. 200, 1527–1536.
Chai, P., Altshuler, D.L., Stephens, D.B., Dillon, M.E., 1999. Maximal horizontal flight performance of hummingbirds: effects of body mass and molt. Physiol. Biochem. Zool. 72, 145–155.
Cornelius, J.M., Perfito, N., Zann, R., Breuner, C.W., Hahn, T., 2011. Physiological trade-offs in self-maintenance: plumage molt and stress physiology in birds. J. Exp. Biol. 214, 2768–2777.
Covas, L., Senar, J.C., Roqué, L., Quesada, J., 2017. Records of fatal attacks by Rose-ringed Parakeets Psittacula krameri on native avifauna. Rev. Cat. Orn. 33, 45–49.
Cramp, S., Perrins, C.M., 1994. The Birds of the Western Palearctic, vol. 8. Oxford University Press, Oxford.
Dawson, A., 2003. A detailed analysis of primary feather moult in the Common Starling Sturnus vulgaris – new feather mass increases at a constant rate. Ibis 145, 335–340.
Dwight, K.L., 1900. The sequence of plumages and moults of the passerine birds of New York. Ann. N. Y. Acad. Sci. 13, 73–360.
Edwards, A.E., Rohwer, S., 2005. Large-scale patterns of molt activation in the flight feathers of two albatross species. Condor 107, 835–848.
Griggio, M., Serra, L., Licheri, D., Campomori, C., Pilastro, A., 2009. Moult speed affects structural feather ornaments in the blue tit. J. Evol. Biol. 22, 782–792.
Guallar, S., Carrillo-Ortiz, J., Quesada, J., 2021. Marginal covert moult in the house sparrow Passer domesticus. Ring. Migr. 36, 95–104.
Guallar, S., Quesada, J., 2023. Data from: moult intensity constraints along the complete moult sequence of the House Sparrow Passer domesticus. Figshare. .
Hall, K.S.S., Fransson, T., 2000. Lesser Whitethroats under time-constraint moult more rapidly and grow shorter wing feathers. J. Avian Biol. 31, 583–587.
Haukioja, E., 1971. Flightlessness in some moulting passerines in northern Europe. Ornis Fenn. 48, 101–116.
Hedenström, A., 1998. The relationship between wing area and raggedness during molt in the willow warbler and other passerines. J. Field Ornithol. 69, 103–108.
Hedenström, A., Sunada, S., 1999. On the aerodynamics of moult gaps in birds. J. Exp. Biol. 202, 67–76.
Hegelbach, J., 2014. Die Mauser der Wasseramsel Cinclus cinclus und der Bezug zu Geschlecht, Alter und Bruttermin. Vogelwarte 52, 179–190.
Jenni, L., Ganz, K., Milanesi, P., Winkler, R., 2020. Determinants and constraints of feather growth. PLoS One 15, e0231925.
Jenni, L., Winkler, R., 2020a. Molt and Ageing of European Passerines, second ed. Bloomsbury, London.
Jenni, L., Winkler, R., 2020b. The Biology of Moult in Birds. Helm, London.
Kasparek, M., 1979. Zum Ablauf der postnuptialen Vollmauser der Rohrammer (Emberiza schoeniclus). J. Ornithol. 120, 247–264.
Kiat, Y., 2017. Divergent primary moult: a rare moult sequence among Western Palaearctic passerines. PLoS One 12, e0187282.
Laesser, J., van Wijk, R., 2018. Postponed moult of primary coverts untangles the ageing of Wrynecks Jynx torquilla. Ringing Migr. 32, 87–103.
Michener, H., Michener, J.R., 1940. The molt of house finches of the Pasadena region. Condor 42, 140–153.
Morton, M.L., Welton, D.E., 1973. Postnuptial molt and its relation to reproductive cycle and body weight in mountain white-crowned sparrows (Zonotrichia leucophrys oriantha). Condor 75, 184–189.
Newton, I., 1966. The moult of the bullfinch Pyrrhula pyrrhula. Ibis 108, 41–67.
Newton, I., 1967. Feather growth and moult in some captive finches. Hous. Theor. Soc. 14, 10–24.
Noordhuis, R., 1989. Patronen in slagpenrui oecofysiologische aanpassingen. Limosa 62, 35–45.
Pyle, P., 2005. Molts and plumages of ducks (Anatinae). Waterbirds 28, 208–219.
Pyle, P., Howell, S.N.G., 1995. Flight feather molt patterns and age of North American woodpeckers. J. Field Ornithol. 66, 564–581.
Pyle, P., Tranquillo, K., Kayano, K., Arcilla, N., 2016. Molt patterns, age criteria, and molt-breeding dynamics in American Samoan landbirds. Wilson J. Ornithol. 128, 56–69.
R Core Team, 2022. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. .
Randall, D., Burggren, W., French, K., 2001. Eckert Animal Physiology, fifth ed. Freeman, New York.
Rohwer, S., 2008. A primer on summarizing molt data for flight feathers. Condor 110, 799–806.
Rohwer, S., Broms, K., 2013. Replacement rules for the flight feathers of yellow-billed cuckoos (Coccyzus americanus) and common cuckoos (Cuculus canorus). Auk 130, 599−608.
Rohwer, S., Ricklefs, R.E., Rohwer, V.G., Copple, M.M., 2009. Allometry of the duration of flight feather molt in birds. PLoS Biol. 7, e1000132.
Sach, G., 1968. Die Mauser des Großen Brachvogels, Numenius arquata. J. Ornithol. 109, 485–511.
Salewski, V., Bairlein, F., Leisler, B., 2002. Remige moult in Spotted Flycatcher (Muscicapa striata) on its West African wintering grounds. Vogelwarte 41, 301–303.
Seel, D.C., 1976. Moult in five species of Corvidae in Britain. Ibis 118, 491–536.
Stekhoven, D.J., Buehlmann, P., 2012. MissForest – non-parametric missing value imputation for mixed-type data. Bioinformatics 28, 112–118.
Swaddle, J.P., Witter, M.S., 1997. The effects of molt on the flight performance, body mass, and behavior of European starlings (Sturnus vulgaris): an experimental approach. Can. J. Zool. 75, 1135–1146.
Vágási, C.I., Pap, P.L., Barta, Z., 2010. Haste makes waste: accelerated molt adversely affects the expression of melanin-based and depigmented plumage ornaments in House Sparrows. PLoS One 5, e14215.
Vágási, C.I., Pap, P.L., Vincze, O., Benkő, Z., Marton, A., Barta, Z., 2012. Haste makes waste but condition matters: molt rate-feather quality trade-off in a sedentary songbird. PLoS One 7, e40651.
van Zeeland, Y.R., Schoemaker, N.J., 2014. Plumage disorders in psittacine birds-part 1: feather abnormalities. Eur. J. Comp. Anim. Pract. 24, 34–47.
Wijnandts, H., 1984. Ecological energetics of the Long-Eared Owl (Asio otus). Ardea 72, 1–92.
Williams, E.V., Swaddle, J.P., 2003. Moult, flight performance and wingbeat kinematics during take-off in European starlings Sturnus vulgaris. J. Avian Biol. 34, 371–378.
Williamson, K., 1961. Sequence of the post-nuptial moult in the starling. Bird Migration 2, 43–45.
Williamson, K., 1972. Reversal of normal moult sequence in the Spotted Flycatcher. Br. Birds 65, 50–51.
Wingfield, J.C., Lynn, S.E., Soma, K.K., 2001. Avoiding the 'costs' of testosterone: ecological bases of hormone-behavior interactions. Brain Behav. Evol. 57, 239–251.
Winkler, R., Daunicht, W.D., Underhill, L.G., 1988. Die Grossgefiedermauser von Alpendohle Pyrrhocorax graculus und Alpenkrähe Pyrrhocorax pyrrhocorax. Ornithol. Beob. 85, 245–259.
Zeidler, K., 1966. Untersuchungen über Flügelbefiederung und Mauser des Haussperling Passer domesticus. J. Ornithol. 107, 113–115.
Zuberogoitia, I., Zabala, J., Martínez, J.E., 2018. Moult in birds of prey: a review of current knowledge and future challenges for research. ARDEOLA 65, 183–207.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds