Juan A. Amat, Nico Varo, Marta I. Sánchez, Andy J. Green, Dámaso Hornero-Méndez, Juan Garrido-Fernández, Cristina Ramo. 2023: Physiological strategies of moult-migrating Black-necked Grebes (Podiceps nigricollis) in a polluted staging site according to blood chemistry. Avian Research, 14(1): 100118. DOI: 10.1016/j.avrs.2023.100118
Citation:
Juan A. Amat, Nico Varo, Marta I. Sánchez, Andy J. Green, Dámaso Hornero-Méndez, Juan Garrido-Fernández, Cristina Ramo. 2023: Physiological strategies of moult-migrating Black-necked Grebes (Podiceps nigricollis) in a polluted staging site according to blood chemistry. Avian Research, 14(1): 100118. DOI: 10.1016/j.avrs.2023.100118
Juan A. Amat, Nico Varo, Marta I. Sánchez, Andy J. Green, Dámaso Hornero-Méndez, Juan Garrido-Fernández, Cristina Ramo. 2023: Physiological strategies of moult-migrating Black-necked Grebes (Podiceps nigricollis) in a polluted staging site according to blood chemistry. Avian Research, 14(1): 100118. DOI: 10.1016/j.avrs.2023.100118
Citation:
Juan A. Amat, Nico Varo, Marta I. Sánchez, Andy J. Green, Dámaso Hornero-Méndez, Juan Garrido-Fernández, Cristina Ramo. 2023: Physiological strategies of moult-migrating Black-necked Grebes (Podiceps nigricollis) in a polluted staging site according to blood chemistry. Avian Research, 14(1): 100118. DOI: 10.1016/j.avrs.2023.100118
After breeding, Black-necked Grebes (Podiceps nigricollis) perform a moult-migration to autumn hypersaline staging sites, where they moult the flight feathers and forage on superabundant brine shrimp (Artemia spp.) before leaving for wintering areas. During the stay in moulting sites, the grebes experience changes in organs and muscle size (atrophy, hypertrophy), and almost double their body mass, which has been suggested to act as an insurance against a collapse in prey availability in late autumn. During two years we collected blood samples from hundreds of individuals at one of the most important European moulting sites (the Odiel marshes, SW Spain), which is a highly polluted area due to mining drainage and chemical industry. We assessed the potential effect of moulting stage, day of the year and body condition on 16 blood biochemical parameters. Because of the changes in prey availability and body composition of grebes, we expected some physiological adjustments during moult. Elevated levels of cholesterol suggested that birds in active moult increased foraging effort to face the costs of moulting. There was increased amount of lactate dehydrogenase, corresponding to periods of breast muscle atrophy. Birds in active moult augmented protein ingestion, likely to account for the requirements of feather growth. We also show that the probability of fasting due to low prey availability increased late in the moulting season, as demonstrated by an increase in plasma β-hydroxy-butyrate.
Several bird species that simultaneously moult their flight feathers must rely on body reserves or on abundant food during feather replacement (Hohman et al., 1992; Moorman et al., 1993), and in many waterbirds this is accomplished by performing a moult migration to specific sites following breeding (Salomonsen, 1968; Jehl Jr, 1990). One of such species is the Black-necked Grebe (or Eared Grebe, Podiceps nigricollis), which in late summer and autumn congregates in saline wetlands during staging. The most important of these sites are hypersaline, where the superabundance of brine shrimp Artemia spp. facilitates the aggregation of thousands of individuals to moult (Cooper et al., 1984; Jehl Jr, 1988; Varo et al., 2011). The abundance of brine shrimps in these wetlands decreases dramatically in late autumn (Cooper et al., 1984; Varo et al., 2011) and grebes then leave the moulting staging sites for wintering areas (Storer and Jehl Jr, 1985; Jehl Jr, 1988; Varo et al., 2011). Jehl (1997) reported differences in the body composition of Black-necked Grebes due to changes in muscle and organs size, as well as in fat deposition during staging in moulting sites. During fattening, the grebes may almost double their body mass (Jehl Jr, 1997). It has been suggested that such large fat reserves act as an insurance against a population collapse of brine shrimps in late autumn (Jehl Jr, 1988). However, depending on the date of moulting, not all individuals adopted the same strategy of fat deposition in Spain. While early-moulting grebes accumulated fat stores during moulting and were lighter than late-moulting birds, delayed moulters acquired greater fat stores in advance of moult and retained extra stores later, most likely as a bet hedge against the increasing probability of food supply failing late in the season (Fox et al., 2013).
Because of the changes in body composition of Black-necked Grebes, some physiological adjustments during moult are expected (Jenni and Winkler, 2020). In Black-necked Grebes moulting in Mono Lake (California, USA), Yoshida (2017) found a positive correlation between body mass and both triglycerides and uric acid in plasma, indicative of fat deposition and protein catabolism, respectively. In contrast, some plasma biochemical parameters do not show the same pattern in other moulting waterbirds (Podlaszczuk et al., 2017), which may be due to differences between species in their moulting ecology strategies (Ghebremeskel et al., 1989).
The Odiel marshes, where we conducted our study, is a highly polluted area in the joint estuary of the Tinto and Odiel rivers in South-West Spain (Sainz et al., 2004; Pérez-López et al., 2011; Vallés et al., 2017). The origin of this pollution is the acid mine drainage from historic mining activities in the Iberian Pyrite Belt, a massive polymetallic sulphide deposit (Nocete et al., 2005). Pollution also derives from effluents of the chemical industry at the mouth of the Tinto and Odiel rivers, resulting in high levels of As, Cd, Cu, Pb and Zn (Sarmiento et al., 2009). Brine Shrimp (Artemia parthenogenetica) may be the main route of metal exposure for grebes in the Odiel marshes (Sánchez et al., 2013; Rodríguez-Estival et al., 2019), as pollutants accumulate in their invertebrate prey (Pais-Costa et al., 2021). Indeed, a study at this site found potentially toxic levels of As, Hg and Zn in Black-necked Grebes (Rodríguez-Estival et al., 2019). These elements may affect the metabolism and immune responses in animals (Kidd et al., 1996; Hoffman and Heinz, 1998; Suzuki et al., 2004; Wu et al., 2022). In particular, in the Common Loon Gavia immer and the Red-necked Grebe Podiceps grisegena lipid reserves were negatively correlated with Hg concentrations (Evers et al., 2008; Holman et al., 2015), and in the Greater Flamingo Phoenicopterus roseus at our study site there was an adverse effect of Hg bioaccumulation on body condition (Gillingham et al., 2021).
In this study we analysed several blood chemistry parameters in Black-necked Grebes during their flight feather moult at the Odiel marshes. First, we investigated how blood parameters inform on the physiological changes that Black-necked Grebes experience during the moult of flight feathers. And second, we expected that if grebes experience fasting during the late staging period because of the population collapse of its main Artemia prey (Varo et al., 2011), this could be detected in their plasma metabolites, particularly β-hydroxy-butyrate, which is involved in fat catabolism and increases in concentration during fasting (Jenni-Eiermann and Jenni, 1994; Williams et al., 1999).
2.
Materials and methods
2.1
Study site and sample collection
This study was carried out at the Odiel marshes in southwestern Spain (37°14ʹ N, 6°57ʹ W). Part of the marshes was transformed in a saltwork in 1974, in which Black-necked Grebes use the evaporation ponds for foraging (Sánchez et al., 2006). During mid-summer and late-autumn the marshes are used as a moulting staging site by thousands of grebes from Central Europe and South-Central Russia (Varo et al., 2011; Amat et al., 2014).
Using procedures described in Amat et al. (2014) and Sáez-Gómez et al. (2017), grebes were captured for ringing purposes every 1–2 weeks from late August until early December in 2008 and 2009. Based on iris color, the grebes were classified as birds of the year or adults, with adults having redder irises (Storer and Jehl Jr, 1985). As birds of the year do not moult flight feathers in autumn staging sites (Cullen et al., 2020), only adult birds were considered here. The moulting stage of every individual was allocated to one of three categories: unmoulted, moulting and moulted (Fox et al., 2013). The grebes were: (1) visually sexed by an experienced ringer, who assigned sex correctly to 85% of the birds, as corroborated using molecular procedures (Amat et al., 2014); (2) measured (head-bill length) to the nearest mm; and (3) weighed to the nearest 5 g. We estimated body condition according to the Scaled Mass Index proposed by Peig and Green (2009) following procedures in Rodríguez-Estival et al. (2019). The index was standardized to a mean head-bill length of 56.91 mm.
We collected 1 mL of blood from the tarsal vein in heparinized tubes (388 samples in 2008, and 279 in 2009). Blood collection was usually done within 2 h following capture, at around 13:00 ± 1 h (GMT). As this time of day encompasses the daily feeding period of grebes (Varo et al., 2011), the blood parameters in our samples were not affected by the eat-fast daily cycle (Ferrer et al., 1994). The tubes were kept at 4 ℃ until transportation to a laboratory where the blood samples were centrifuged (10 min at 4000 rpm, corresponding to a relative centrifugal force of 2688 g) to separate the plasma, which was stored in cryovials at −80 ℃ until analyses. The separation of plasma was done less than 5 h after the birds were bled. The analyses of all blood parameters considered (Table 1) were carried out in a Cobas Integra® 400 (Roche, Germany), a fully automated, computerized chemistry analyser. The blood samples were also used for molecular sexing, following procedures in Amat et al. (2014) and Sáez-Gómez et al. (2017), though not all individuals were molecularly sexed due to budget constraints.
Table
1.
Median values (with values of the 25th and 75th percentiles in parentheses) of plasma chemistry parameters from adult Black-necked Grebes according to moulting stage (sample sizes in parentheses).
Parameter
Unmoulted (62)
Moulting (160)
Moulted (445)
χ22
Albumin (g/dL)
1.35 (1.27–1.50)
1.40 (1.27–1.51)
1.45 (1.35–1.56)
20.82***
Alkaline phosphatase (U/L)
19.79 (14.27–27.83)
23.72 (18.01–29.79)
18.04 (14.32–21.88)
55.04***
β-hydroxy-butyrate (μmol/L)
1051.44 (751.35–1557.97)
1247.53 (800.20–1751.45)
1338.71 (896.88–1959.76)
8.44*
Calcium (mg/dL)
10.34 (9.70–11.21)
10.50 (10.03–11.08)
10.40 (9.96–10.94)
3.70ns
Cholesterol (mg/dL)
320.97 (268.82–374.08)
295.715 (259.30–326.74)
370.76 (306.98–424.53)
93.75***
Creatine kinase (U/L)
1650.50 (1212.50–2247.50)
1407.00 (1136.75–1907.75)
1577.00 (1172.00–2276.00)
8.32*
Glucose (mg/dL)
325.49 (290.41–344.77)
294.07 (267.51–327.29)
324.90 (294.80–352.56)
40.29***
Lactate dehydrogenase (U/L)
366.00 (262.25–537.00)
382.00 (284.00–520.25)
364.00 (252.00–560.00)
0.58ns
Magnesium (mmol/L)
0.94 (0.86–1.06)
0.99 (0.92–1.00)
0.92 (0.85–1.02)
29.99***
Phosphorus (mg/dL)
1.76 (1.31–2.57)
2.64 (1.89–3.16)
2.13 (1.63–2.64)
40.13***
Potassium (mmol/L)
1.78 (1.22–2.57)
2.04 (1.53–2.51)
1.78 (1.33–2.55)
7.95*
Sodium (mmol/L)
151.35 (146.80–158.28)
151.20 (146.30–156.98)
152.20 (147.70–158.30)
2.37ns
Total proteins (g/dL)
3.80 (3.43–4.19)
3.98 (3.47–4.52)
3.84 (3.48–4.21)
6.64*
Triglycerides (mg/dL)
147.42 (120.47–191.17)
157.75 (136.69–176.66)
178.49 (146.89–207.69)
34.67***
Urea (mg/dL)
5.88 (4.90–7.03)
4.67 (3.79–5.79)
6.54 (5.08–8.19)
80.34***
Uric acid (mg/dL)
8.14 (7.17–9.20)
6.59 (5.38–8.07)
7.56 (5.80–9.51)
22.96***
Differences between moulting stages were tested using Kruskall-Wallis ANOVA (χ22). ns: non-significant; *p < 0.05; ***p < 0.001.
These were conducted using jamovi 2.3.13 (The jamovi project, 2022). Although some individuals were captured multiple times, we only used the first capture of every individual for statistical purposes, as many of the recaptures were within one or two weeks after the first capture (i.e., the grebes were in the same moulting stage). Data for blood chemistry values and body measurements of grebes used in this study, including individual identity (ring number) and sampling dates, are available in Amat et al. (2023).
We used principal component analysis (PCA) to reduce the complexity in the dataset, applying Varimax rotation. For the PCA the values of all blood parameters were log-transformed. The number of interpretable ordination axes was determined by the statistical programme using the scree plot method, and we determined the significance of loadings using the broken-stick criterion (Peres-Neto et al., 2003). Generalised Linear Mixed Models (GLMMs) were performed to examine relationships between PC scores and body condition (log-transformed), date of sample collection (day of the year) and moulting stage. In these models the PC scores were the dependent variables, whereas moult stage was a fixed factor, and date of sampling and body condition were continuous variables. Year was considered as a random factor in the GLMMs. It is recommended that at least five levels of random effects should be used in GLMMs, otherwise the variance estimates of random effects may be biased (Silk et al., 2020). However, we were not interested in between-year variation after controlling for repeated measures, and under these circumstances two levels of random effects may be used (Gomes, 2021). Sex was not included in the final models because there were no sex-related differences in blood parameters. We used post-hoc comparisons to assess differences between moulting stages, adjusting probabilities by the Bonferroni method.
Spearman rank-correlation was used to examine the relationship between β-hydroxy-butyrate concentration and date of sampling.
3.
Results
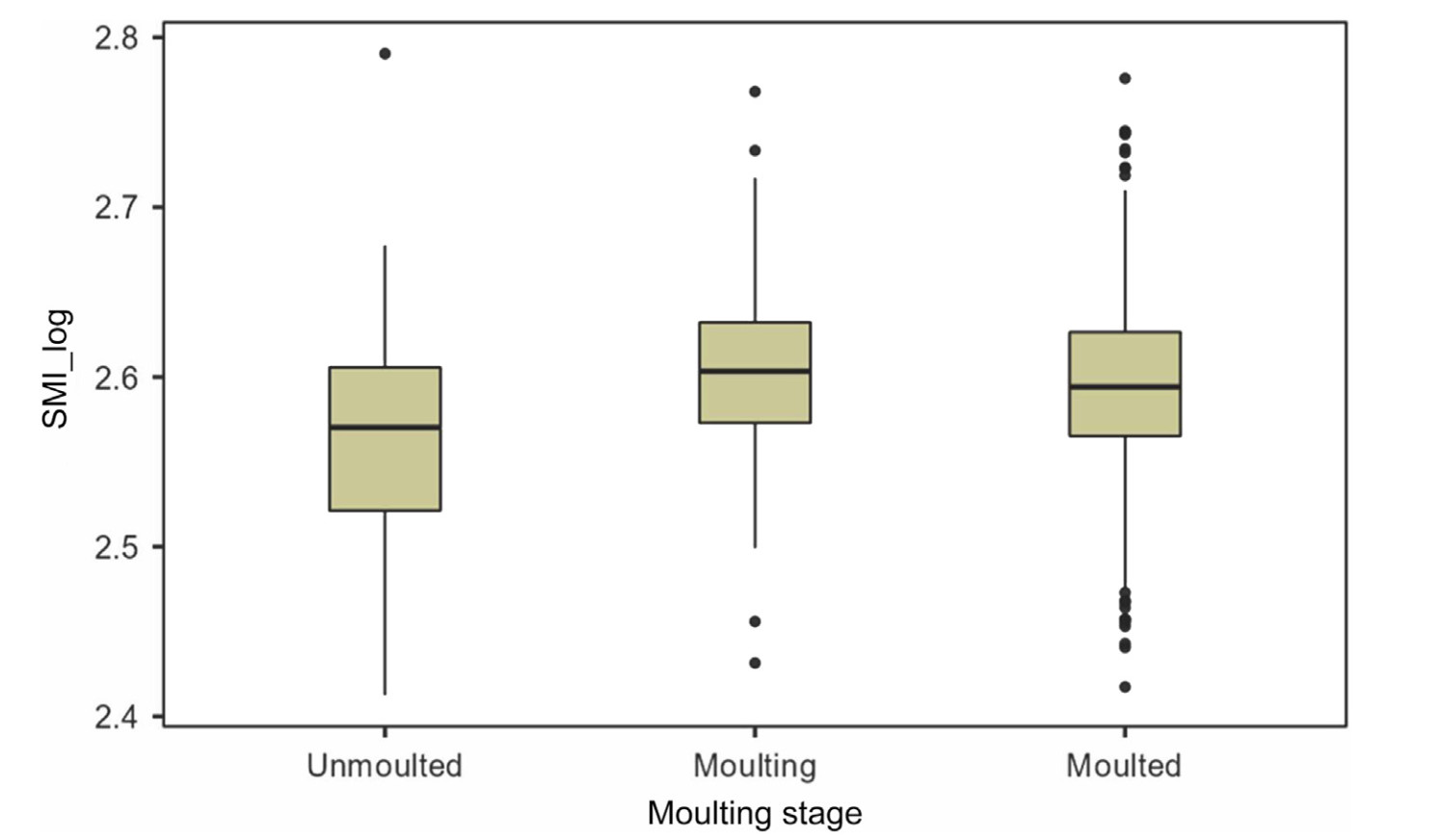
The concentrations of all blood parameters, except calcium, lactate dehydrogenase, and sodium, varied with the moulting stage (Table 1). β-hydroxy-butyrate was higher in individuals that had completed moult than in individuals in other moulting stages, and there was a highly significant correlation between β-hydroxy-butyrate levels and date of sampling (rS = 0.37, n = 667, p < 0.001). There were variations in body condition depending on moulting stage, the grebes being in higher condition when they were in active moult (one-way ANOVA, F2,664 = 9.67, p < 0.001; Fig. 1).
Figure
1.
Boxplots for body condition (scaled mass index, log-transformed) of Black-necked Grebes according to moulting stage. The boxes represent the interquartile ranges (IQR: 25th and 75th percentiles), and the lines within the boxes are the medians. Whiskers extend 1.5 × IQR from the 75th and 25th percentiles. The dots denote outliers.
The first four principal axes of a PCA on the blood values explained 51.7% of the variance (Table 2). The first principal axis (PC1) was positively associated with cholesterol (Table 2). Albumin and total proteins were positively associated with PC2, and lactate dehydrogenase was positively associated with PC3 (Table 2). No parameter loaded significantly on PC4, though phosphorus and potassium loaded positively and uric acid negatively, all with moderate values (Table 2).
Table
2.
Results from Principal Component Analysis (using Varimax rotation) of plasma chemistry parameters (log-transformed) of adult Black-necked Grebes.
Parameter
PC1
PC2
PC3
PC4
Albumin
0.494
0.726
−0.210
−0.015
Alkaline phosphatase
0.070
0.094
−0.373
−0.239
β-hydroxy-butyrate
0.345
0.013
−0.097
0.463
Calcium
0.034
0.680
0.028
0.020
Cholesterol
0.848
0.226
−0.100
−0.043
Creatine kinase
0.244
−0.046
0.673
0.094
Glucose
0.388
0.124
−0.473
0.017
Lactate dehydrogenase
−0.026
−0.048
0.816
−0.114
Magnesium
−0.339
0.473
0.523
−0.298
Phosphorus
−0.125
0.200
0.350
0.609
Potassium
0.085
−0.101
0.091
0.642
Sodium
0.203
0.414
−0.094
−0.035
Total proteins
0.007
0.809
−0.052
−0.035
Triglycerides
0.718
0.184
−0.019
−0.052
Urea
0.550
−0.084
0.385
0.042
Uric acid
0.277
0.087
0.187
−0.603
% variance explained
14.8
13.9
13.4
9.6
Values for every parameter show loadings of the first four principal component (PC) axes. Values in bold are significant according to the brocken-stick criterion (Peres-Neto et al., 2003).
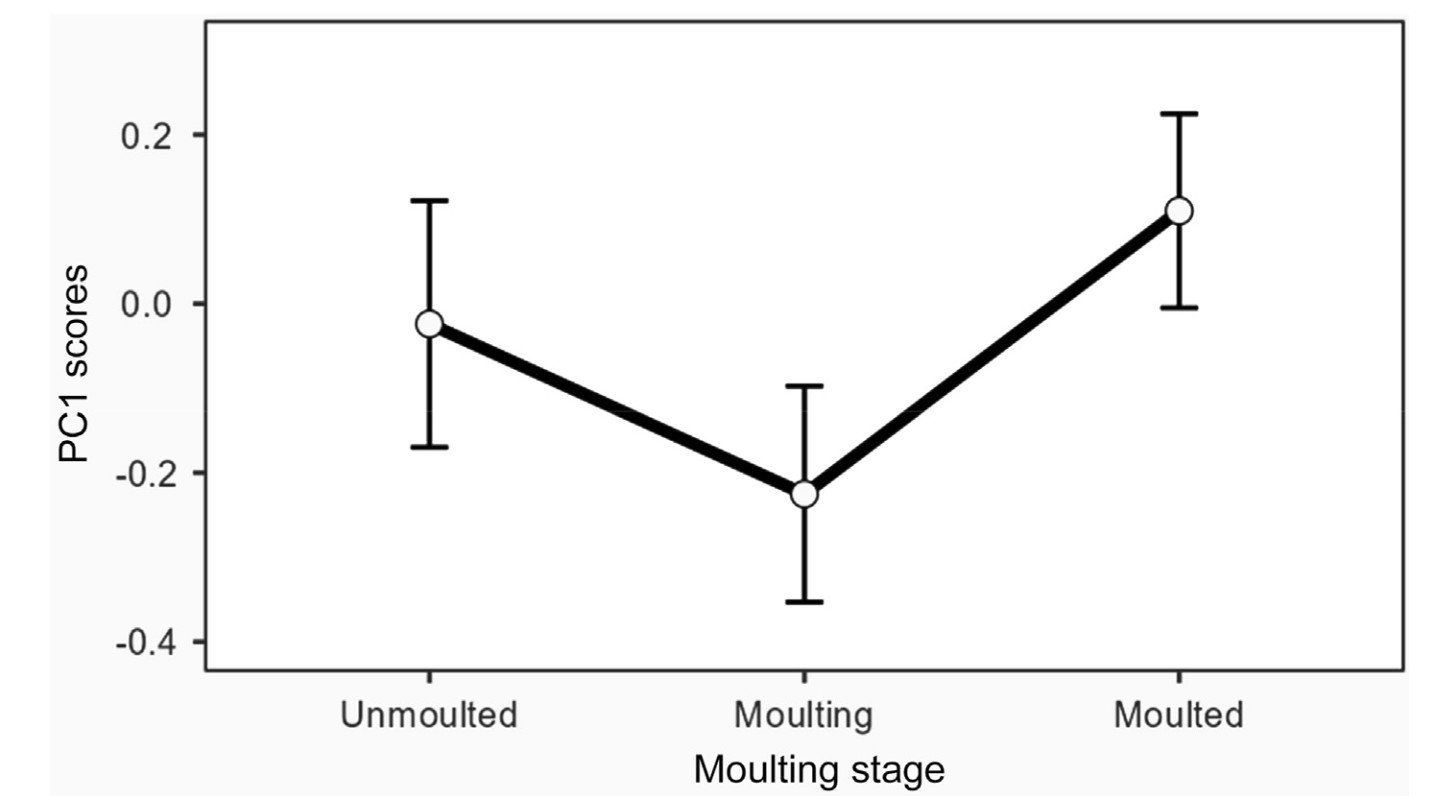
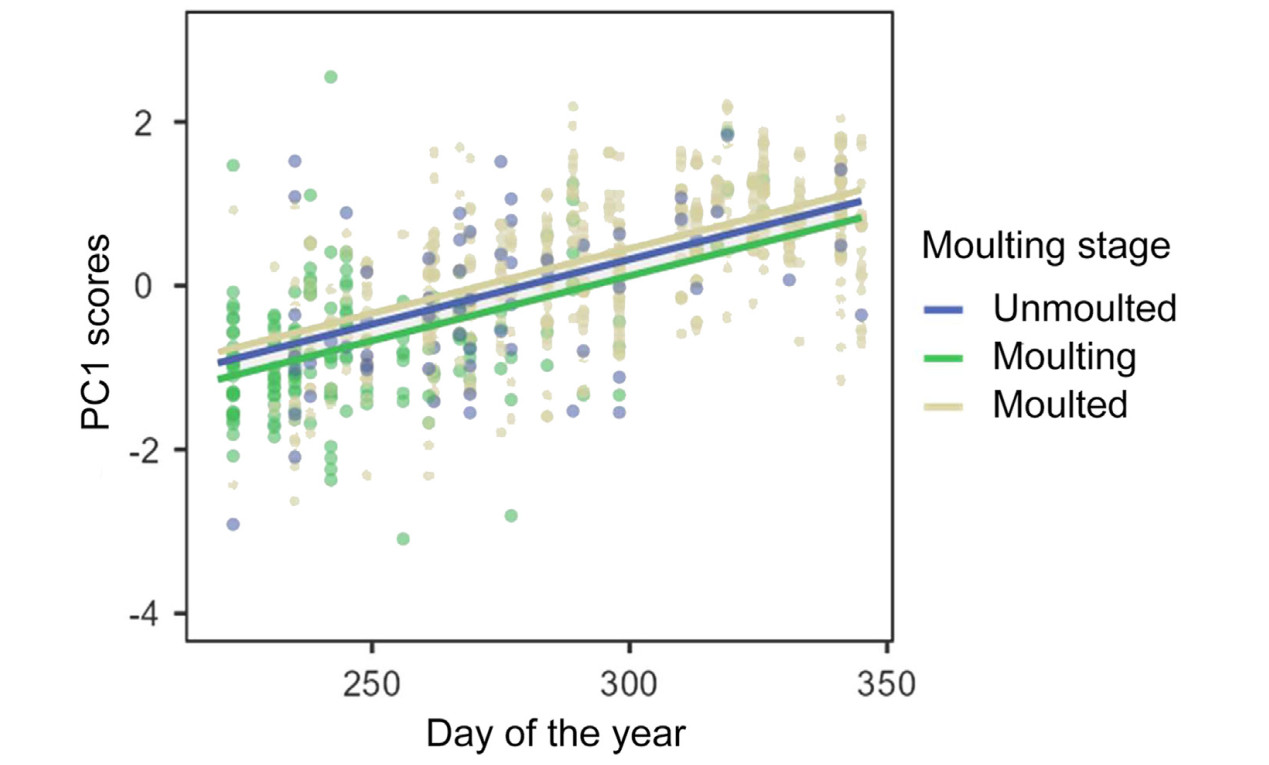
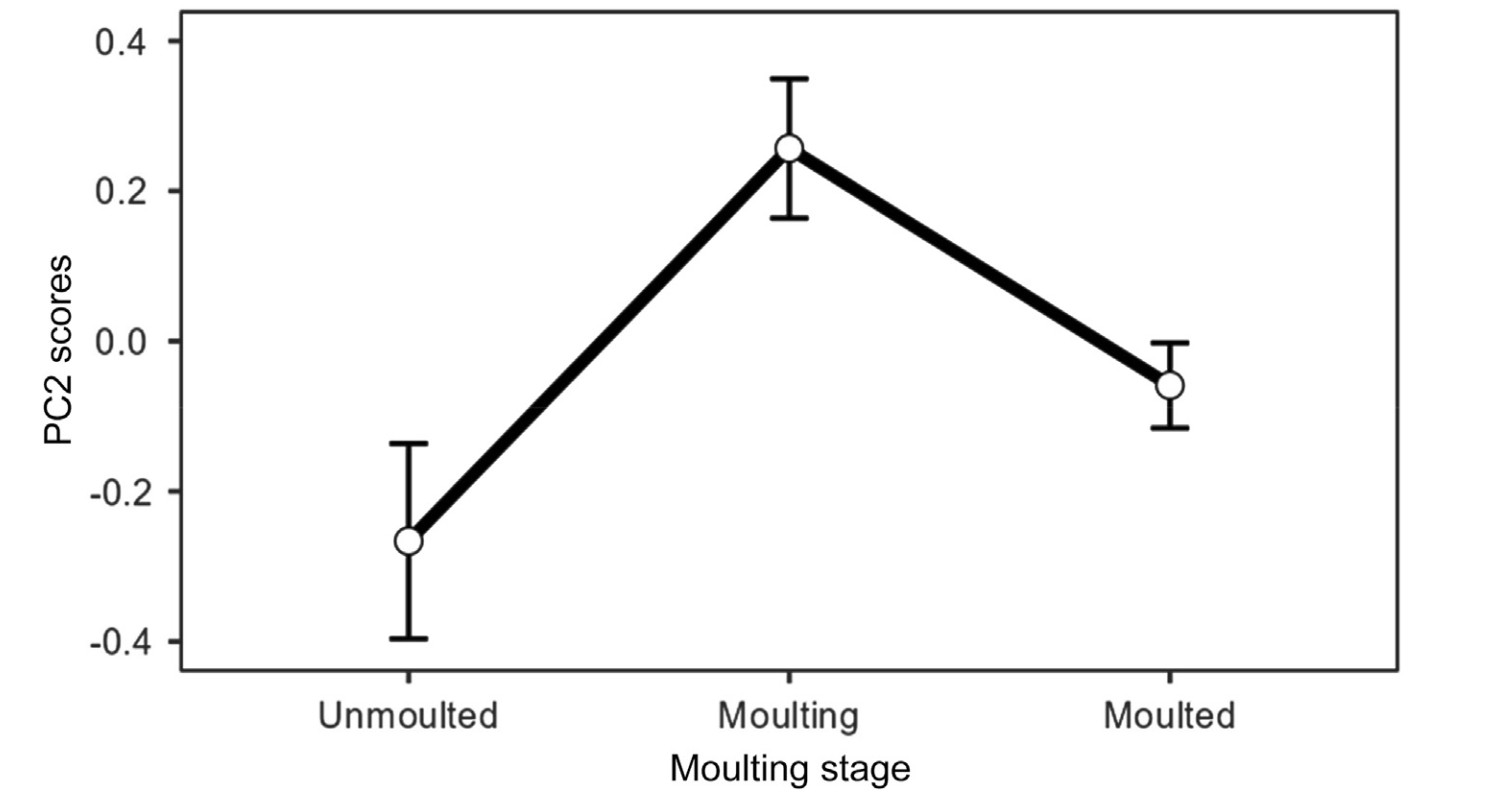
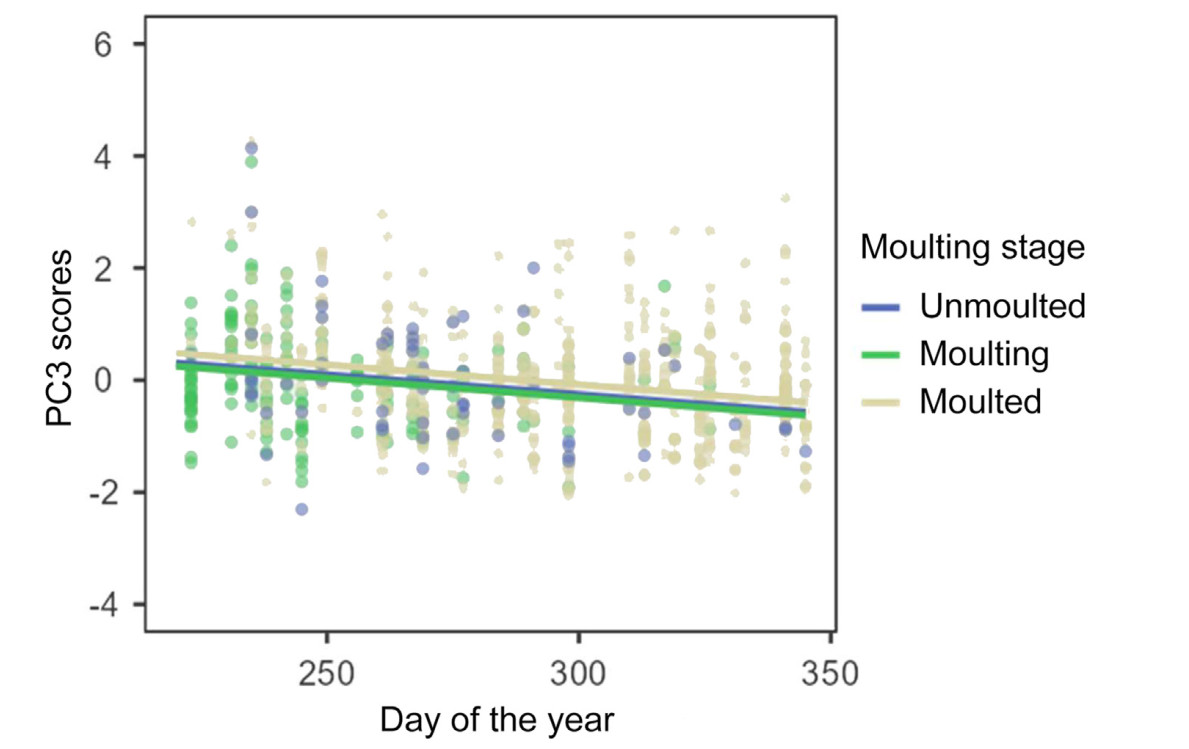
Of the scores of the first three axes (i.e., those on which some variable loaded significantly), those of PC1 were affected by moulting stage and date (Table 3). Differences in the PC1 scores were only significant between moulting and moulted grebes (difference = −0.34 ± (s.e.) 0.08; t = −4.01, p < 0.001) (Fig. 2). There was an increase of PC1 scores throughout the staging season (Fig. 3). PC2 scores were only affected by moulting stage, (Table 3), being higher for moulting birds than for those unmoulted (difference = 0.52 ± 0.16; t = 3.35, p = 0.003), as well as for grebes that had completed moult (difference = 0.32 ± 0.11; t = 2.86, p = 0.013) (Fig. 4). Finally, PC3 scores were significantly affected only by date of sampling (Table 3), with a decrease of PC3 scores throughout the season (Fig. 5).
Table
3.
Generalised linear mixed models (GLMMs) of PC scores of adult Black-necked Grebes in relation to moulting stage (unmoulted, moulting, and moulted), date of capture and body condition (log-transformed scaled body mass).
Source of variation
F
Num df
Den df
P
PC1 scores
Moulting stage
8.12
2
660.7
< 0.001
Body condition
2.88
1
660.7
0.090
Date
268.38
1
660.5
< 0.001
PC2 scores
Moulting stage
6.79
2
649.8
0.001
Body condition
0.70
1
651.4
0.403
Date
1.18
1
658.8
0.279
PC3 scores
Moulting stage
2.76
2
660.8
0.064
Body condition
0.32
1
660.8
0.572
Date
31.63
1
660.5
< 0.001
The scores resulted from a Principal Component Analysis of plasma chemistry parameters. Numerator (Num) and Denominator (Den) of the df for the F values are given.
Figure
2.
Predicted patterns of variation (means ± s.e.) showing PC1 scores according to moulting stages of adult Black-necked Grebes. Mean and s.e. values stem from a GLMM in Table 3. The scores resulted from a Principal Component Analysis of plasma chemistry parameters from adult grebes.
Figure
3.
Relationship between PC1 scores and date of sampling (day of year) according to moulting stage of Black-necked Grebes. The regression lines stem from a GLMM in Table 3. The scores resulted from a Principal Component Analysis of plasma chemistry parameters from adult grebes.
Figure
4.
Predicted patterns of variation (means ± s.e.) showing PC2 scores according to moulting stages of adult Black-necked Grebes. Mean and s.e. values stem from a GLMM in Table 3. The scores resulted from a Principal Component Analysis of plasma chemistry parameters from adult grebes.
Figure
5.
Relationship between PC3 scores and date of sampling (day of year) according to moulting stage of Black-necked Grebes. The regression lines stem from a GLMM in Table 3. The scores resulted from a Principal Component Analysis of plasma chemistry parameters from adult grebes.
We found evidence that there were temporal-related variations in the physiology of Black-necked Grebes during the moulting period in autumn, as revealed by the analysis of several chemistry parameters in plasma. Because of the time elapsed between stopping feeding and blood sampling, the blood parameters may be biased towards fasting. However, birds were similarly affected by this potential problem throughout the season, so that the differences between grebes in different moulting stages are still valid, assuming that the time interval between capture and blood sampling was similar among groups.
There were some differences in the blood parameters that loaded significantly on PC axes that varied across the moulting season. Cholesterol loaded significantly on PC1. This lipid is considered a marker of nutritional status (Alonso-Alvarez et al., 2003; Amat et al., 2007), and an increase in dietary cholesterol would indicate that the grebes were gaining body mass (Alonso-Alvarez et al., 2002). PC1 scores were affected by date, being greater as the season progressed, which likely was due to increased foraging effort of the grebes throughout the season (Varo et al., 2011). In addition to date, PC1 scores were also affected by moulting stage, being greater in grebes that had moulted, which may indicate that these birds were gaining mass as the season progressed to face the collapse in prey availability late in the season (see above).
PC2 was associated with albumin and total proteins, and was affected by moulting stage, PC2 scores being lower in unmoulted and moulted birds than in grebes in active moult. The association of this PC axis with albumin indicates an exhaustion of lipid reserves after the moult migration to Odiel marshes and also that some birds could be using lipid reserves after moulting (Vleck and Vleck, 2002; see below). The association of PC2 with total proteins is likely an indication of increased protein requirements for feather growth in birds in active moult.
PC3 was associated with lactate dehydrogenase. This enzyme has been used as a diagnostic indicator of muscle fitness (Knuth and Chaplin, 1994). In moulting Black-necked Grebes an increase in heart and leg masses and atrophy of breast muscles was found during the flightless moult in North America, but the changes were reversed when the grebes prepared to depart for wintering areas (Jehl Jr, 1997). PC3 scores were negatively associated with date of sampling, suggesting that lactate dehydrogenase values were affected by the changes in breast muscles mass, in particular with their atrophy. The increase of breast muscles mass when grebes prepare to depart for wintering sites may be facilitated by an increase in flapping activity throughout the moulting season (Varo et al., 2011) and a concomitant reduction in levels of lactate dehydrogenase in plasma.
Fasting conditions were expected given the sudden decrease in food availability at the end of the staging period (Varo et al., 2011). β-hydroxy-butyrate levels were higher in moulted birds in comparison with grebes in other moulting stages (Table 1), suggesting that individuals that moulted later could be fasting, which is supported by a correlation between β-hydroxy-butyrate levels and date of sampling. Since β-hydroxy-butyrate is involved in the catabolism of fat, this is in agreement with the idea that grebes use fat stores late in the staging period as an insurance against falling food supply (Jehl Jr, 1988; Fox et al., 2013). This would allow late moulting individuals to regrow the breast muscles despite low food availability, which could allow them to regain flying capability in order to move to wintering sites, such as those in North-West Africa. In turn, this would also explain why late moulting birds accumulate more body reserves than early moulting birds (Fox et al., 2013).
The Odiel marshes is a highly polluted site in which no special measures to decrease contamination levels have been implemented since we conducted our study. The effects of such pollution may have negative impacts on the biota (see Introduction). If pollution has an effect on metabolic processes of Black-necked Grebes, the Odiel marshes, which nowadays is one of the most important staging sites for the species in Europe (Varo et al., 2011; Amat et al., 2014), could act as an ecological trap for these birds during moulting. Nevertheless, to determine if pollution affects grebe performance, a detailed study is needed, for instance estimating survival in relation to pollution levels.
5.
Conclusions
Our study is the first one that uses a comprehensive dataset to analyze physiological responses of Black-necked Grebes during autumn moulting. This species is particularly interesting due to the large changes in body mass during moulting and because it experiences a collapse in prey availability late in autumn. This collapse can occur before many individuals have left for wintering sites, thus potentially causing them to become "trapped" in the staging site. We have shown that the probability of fasting increases late in the moulting season, as demonstrated by a concomitant increase in plasma β-hydroxy-butyrate. Likely, greater accumulation of fat of late moulting individuals in relation to earlier moulters serves to withstand such a period of fasting. This temporal increase in fat reserves is accomplished by a temporal increase in food ingestion, as demonstrated by increases in cholesterol throughout the autumn staging period.
Authors' contributions
All authors conceived the study. JAA, NV, AJG and CR contributed to data collection. JAA performed analysis of data and wrote the first draft. All authors contributed to the interpretation of results and writing of the final manuscript. All authors read and approved the final manuscript.
Ethics statement
All work was carried out under the required legal statements. Consejería de Medio Ambiente, Junta de Andalucía, authorised the fieldwork and the taking of samples.
Declaration of competing interest
The authors declare that they have no competing financial interests that may have influenced the work reported in this paper.
Acknowledgements
We thank all volunteers of EBD-CSIC and SEO/BirdLife that, led by Luis García from Equipo de Seguimiento de Procesos Naturales (ICTS-EBD), participated in the captures of grebes. We thank Enrique Martínez (Consejería de Medio Ambiente, Junta de Andalucía), Director of the Paraje Natural Marismas del Odiel, for facilities to conduct fieldwork. Molecular sexing of birds was performed by Mónica Gutiérrez at the Laboratorio de Ecología Molecular (LEM-EBD). Manuel Vázquez collected blood samples and Francisco Miranda analysed them at the Laboratorio de Ecofisiología (LEF-EBD). During writing we benefitted from the facilities of the ICTS-RBD in Doñana. Our thanks also go to two anonymous referees for very helpful and detailed comments.
Alonso-Alvarez, C., Ferrer, M., Viñuela, J., Amat, J.A., 2003. Plasma chemistry of the chinstrap penguin Pygoscelis antarctica during fasting periods: a case of poor adaptation to food deprivation? Polar Biol. 26, 14–19. .
Alonso-Alvarez, C., Velando, A., Ferrer, M., Veira, J.A.R., 2002. Changes in plasma biochemistry and body mass during incubation in the yellow-legged gull. Waterbirds 25, 253–258. .
Amat, J.A., Hortas, F., Arroyo, G.M., Rendón, M.A., Ramírez, J.M., Rendón-Martos, M., et al., 2007. Interannual variations in feeding frequencies and food quality of greater flamingo chicks (Phoenicopterus roseus): Evidence from plasma chemistry and effects on body condition. Comp. Biochem. Physiol. A 147, 569–576. .
Amat, J.A., Varo, N., Sánchez, M.I., Green, A.J., Ramo, C., 2014. Female-biased sex ratio in moulting Black-necked Grebes Podiceps nigricollis in southern Spain. Ardea 102, 207–212. .
Amat, J.A., Varo, N., Sánchez, M.I., Green, A.J., Hornero-Méndez, D., Garrido-Fernández, J., et al., 2023. Blood chemistry values of black-necked grebes Podiceps nigricollis from Odiel marshes (SW Spain) during the moulting period. Digital CSIC. .
Cooper, S.D., Winkler, D.W., Lenz, P.H., 1984. The effect of grebe predation on a brine shrimp population. J. Anim. Ecol. 53, 51–64. .
Cullen, S.A., Jehl Jr., J.R., Nuechterlein, G.L., 2020. In: Billerman, S.M. (Ed.), Eared grebe (Podiceps nigricollis), version 1.0, Birds of the World. Cornell Lab of Ornithology, Ithaca. (Accessed 30 January 2023).
Evers, D.C., Savoy, L.J., de Sorbo, C.R., Yates, D.E., Hanson, W., Taylor, K.M., et al., 2008. Adverse effects from environmental mercury loads on breeding common loons. Ecotoxicology 17, 69–81. .
Ferrer, M., Amat, J.A., Viñuela, J., 1994. Daily variations of blood chemistry values in the chinstrap penguin (Pygoscelis antarctica) during the Antarctic summer. Comp. Biochem. Physiol. 107A, 81–84. .
Fox, A.D., Ramo, C., Varo, N., Sánchez, M.I., Amat, J.A., Green, A.J., 2013. Late-moulting black-necked grebes Podiceps nigricollis show greater body mass in the face of failing food supply. Ibis 155, 814–822. .
Ghebremeskel, K., Williams, G., Keymer, I.F., Horsley, D., Gardner, D.A., 1989. Plasma chemistry of rockhopper (Eudyptes crestatus), magellanic (Spheniscus magellanicus) and gentoo (Pygoscelis papua) wild penguins in relation to moult. Comp. Biochem. Physiol. A 92, 43–47. .
Gillingham, M.A.F., Borghesi, F., Montero, B.K., Migani, F., Béchet, A., Rendón-Martos, M., et al., 2021. Bioaccumulation of trace elements affects chick body condition and gut microbiome in greater flamingos. Sci. Total Environ. 761, 143250. .
Gomes, D.G.E., 2021. Including random effects in statistical models in ecology: Fewer than five levels? BioRxiv. .
Hoffman, D.J., Heinz, G.H., 1998. Effects of mercury and selenium on glutathione metabolism and oxidative stress in mallard ducks. Environ. Toxicol. Chem. 17, 161–166. .
Hohman, W.L., Ankney, C.D., Gordon, D.H., 1992. In: Batts, B.D.J., Afton, A.D., Anderson, M.G., Ankney, C.D., Johnson, D.H., Kadlec, J.A., et al. (Eds.), Ecology and management of postbreeding waterfowl, The Ecology and Management of Breeding Waterfowl. University of Minnesota Press, Minneapolis, pp. 128-189.
Holman, K.L., Schummer, M.L., Petrie, S.A., Chen, Y-W., Belzile, N., 2015. Inorganic contaminants, nutrient reserves and molt intensity in autumn migrant red-necked grebes (Podiceps grisegena) at Georgian Bay. Arch. Environ. Contam. Toxicol. 69, 399–410. .
Jehl Jr., J.R., 1988. Biology of the eared grebe and Wilson's phalarope in the non-breeding season: A study of adaptations to saline lakes. Stud. Avian Biol. 12, 1–74.
Jehl Jr., J.R., 1990. In: Gwinner, E. (Ed.), Aspects of the moult migration, Bird Migration: Physiology and Ecophysiology. Springer, Berlin, pp. 102-113.
Jehl Jr., J.R., 1997. Cyclical changes in body composition in the annual cycle and migration of the eared grebe Podiceps nigricollis. J. Avian Biol. 28, 132–142. .
Jenni, L., Winkler, R., 2020. The Biology of Moult in Birds. Helm, London.
Jenni-Eiermann, S., Jenni, L., 1994. Plasma metabolite levels predict individual body-mass changes in a small long-distance migrant, the garden warbler. Auk 111, 888–899. .
Kidd, M.T., Ferket, P.R., Qureshi, M.A., 1996. Zinc metabolism with special reference to its role in immunity. World Poultry Sci. J. 52, 309–324. .
Knuth, S.T., Chaplin, S.B., 1994. The effect of exercise on plasma activities of lactate dehydrogenase and creatinine kinase in red-tailed hawks (Buteo jamaicensis). J. Raptor Res. 28, 27–33.
Moorman, T.E., Baldassare, G.A., Hess, T.J., 1993. Carcass mass and nutrient dynamics of mottled ducks during remigial molt. J. Wildl. Manag. 57, 224–228. .
Nocete, F., Álex, E., Nieto, J.M., Sáez, R., Bayona, M.R., 2005. An archaeological approach to regional environmental pollution in the south-western Iberian Peninsula related to Third millennium BC mining and metallurgy. J. Archaeol. Sci. 32, 1566–1576. .
Pais-Costa, A.J., Sánchez, M.I., Taggart, M.A., Green, A.J., Hortas, F., Almeida Vinagre, P., et al., 2021. Trace element bioaccumulation in hypersaline ecosystems and implications of a global invasion. Sci. Total Environ. 800, 149349. .
Peig, J., Green, A.J., 2009. New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos 118, 1883–1891. .
Peres-Neto, P.R., Jackson, D.A., Somers, K.M., 2003. Giving meaningful interpretation to ordination axes: assessing loading significance in principal component analysis. Ecology 84, 2347–2363. .
Pérez-López, R., Nieto, J.M., López Cascajosa, M.J., Díaz Blanco, M.J., Sarmiento, A.M., Oliveira, V., et al., 2011. Evaluation of heavy metals and arsenic speciation discharged by the industrial activity on the Tinto-Odiel estuary, SW Spain. Mar. Pollut. Bull. 62, 405–411. .
Podlaszczuk, P., Włodarczyk, R., Janiszewski, T., Kaczmarek, K., Minias, P., 2017. When moult overlaps migration: moult-related changes in plasma biochemistry of migrating common snipe. PeerJ 5, e3057. .
Rodríguez-Estival, J., Sánchez, M.I., Ramo, C., Varo, N., Amat, J.A., Garrido-Fernández, J., et al., 2019. Exposure of black-necked grebes (Podiceps nigricollis) to metal pollution during the moulting period in the Odiel Marshes, Southwest Spain. Chemosphere 216, 774–784. .
Sáez-Gómez, P., Camacho, C., Álvarez, A., Varo, N., Ramo, C., Green, A.J., 2017. Sexual dimorphism and sex determination in black-necked grebes Podiceps nigricollis: on the need to account for geographical variation in morphology and sex ratio. Hous. Theor. Soc. 64, 502–512. .
Sainz, A., Grande, J.A., de la Torre, M.L., 2004. Characterisation of heavy metal discharge into the Ria of Huelva. Environ. Int. 30, 557–566. .
Salomonsen, F., 1968. The moult migration. Wildfowl 19, 5–24.
Sánchez, M.I., Green, A.J., Castellanos, E.M., 2006. Temporal and spatial variation of an aquatic invertebrate community subjected to avian predation at the Odiel salt pans (SW Spain). Arch. Hydrobiol. 166, 199–223. .
Sánchez, M.I., Varo, N., Matesanz, C., Ramo, C., Amat, J.A., Green, A.J., 2013. Cestodes change the isotopic signature of brine shrimp, Artemia, hosts: implications for aquatic food webs. Int. J. Parasitol. 43, 73–80. .
Sarmiento, A.M., Nieto, J.M., Casiot, C., Elbaz-Poulichet, F., Egal, M., 2009. Inorganic arsenic speciation at river basin scales: the Tinto and Odiel rivers in the Iberian Pyrite Belt, SW Spain. Environ. Pollut. 157, 1202–1209. .
Silk, M.J., Harrison, X.A., Hodgson, D.J., 2020. Perils and pitfalls of mixed-effects regression models in biology. PeerJ 8, e9522. .
Storer, R.W., Jehl Jr., J.R., 1985. Moult patterns and moult migration in the black-necked grebe Podiceps nigricollis. Ornis Scand. 16: 253-260. .
Suzuki, N., Yamamoto, M., Watanabe, K., Kambegawa, A., Hattori, A., 2004. Both mercury and cadmium directly influence calcium homeostasis resulting from the suppression of scale bone cells: the scale is a good model for the evaluation of heavy metals in bone metabolism. J. Bone Miner. Metabol. 22, 439–446. .
The jamovi project, 2022. jamovi (version 2.3). (accessed 30 January 2023).
Vallés, S.M., Cambrollé, J., Castillo, J.M., Curado, G., Mancilla-Leytón, J.M., Figueroa-Clemente, E., 2017. In: Finkl, C.W., Makowski, C. (Eds.), Handling high soil trace elements pollution: case study of the Odiel and Tinto Rivers Estuary and the accompanying salt marshes (Southwest Iberian Peninsula), Coastal Wetlands: Alteration and Remediation, vol. 21. Springer International Publishing AG, pp. 215-241. .
Varo, N., Green, A.J., Sánchez, M.I., Ramo, C., Gómez, J., Amat, J.A., 2011. Behavioural and population responses to changing availability of Artemia prey by moulting black-necked grebes, Podiceps nigricollis. Hydrobiologia 664, 163–171. .
Vleck, C., Vleck, D., 2002. Physiological condition and reproduction consequences in Adélie penguins. Integr. Comp. Biol. 42, 76–83. .
Williams, T.D., Guglielmo, C.G., Egeler, O., Martyniuk, C.J., 1999. Plasma lipid metabolites provide information on mass change over several days in captive western sandpipers. Auk 116, 994–1000. .
Wu, H., Wu, R., Chen, X., Geng, H., Hu, Y., Gao, L., et al., 2022. Developmental arsenic exposure induces dysbiosis of gut microbiota and disruption of plasma metabolites in mice. Toxicol. Appl. Pharmacol. 450, 116174. .
Yoshida, A.A., 2017. Plasma Metabolites Reveal Changes in Physiological State of Eared Grebes (Podiceps nigricollis) during Three Non-migratory Periods. Master's Thesis. University of San Diego, San Diego. (Accessed 30 January 2023).
Table
1.
Median values (with values of the 25th and 75th percentiles in parentheses) of plasma chemistry parameters from adult Black-necked Grebes according to moulting stage (sample sizes in parentheses).
Parameter
Unmoulted (62)
Moulting (160)
Moulted (445)
χ22
Albumin (g/dL)
1.35 (1.27–1.50)
1.40 (1.27–1.51)
1.45 (1.35–1.56)
20.82***
Alkaline phosphatase (U/L)
19.79 (14.27–27.83)
23.72 (18.01–29.79)
18.04 (14.32–21.88)
55.04***
β-hydroxy-butyrate (μmol/L)
1051.44 (751.35–1557.97)
1247.53 (800.20–1751.45)
1338.71 (896.88–1959.76)
8.44*
Calcium (mg/dL)
10.34 (9.70–11.21)
10.50 (10.03–11.08)
10.40 (9.96–10.94)
3.70ns
Cholesterol (mg/dL)
320.97 (268.82–374.08)
295.715 (259.30–326.74)
370.76 (306.98–424.53)
93.75***
Creatine kinase (U/L)
1650.50 (1212.50–2247.50)
1407.00 (1136.75–1907.75)
1577.00 (1172.00–2276.00)
8.32*
Glucose (mg/dL)
325.49 (290.41–344.77)
294.07 (267.51–327.29)
324.90 (294.80–352.56)
40.29***
Lactate dehydrogenase (U/L)
366.00 (262.25–537.00)
382.00 (284.00–520.25)
364.00 (252.00–560.00)
0.58ns
Magnesium (mmol/L)
0.94 (0.86–1.06)
0.99 (0.92–1.00)
0.92 (0.85–1.02)
29.99***
Phosphorus (mg/dL)
1.76 (1.31–2.57)
2.64 (1.89–3.16)
2.13 (1.63–2.64)
40.13***
Potassium (mmol/L)
1.78 (1.22–2.57)
2.04 (1.53–2.51)
1.78 (1.33–2.55)
7.95*
Sodium (mmol/L)
151.35 (146.80–158.28)
151.20 (146.30–156.98)
152.20 (147.70–158.30)
2.37ns
Total proteins (g/dL)
3.80 (3.43–4.19)
3.98 (3.47–4.52)
3.84 (3.48–4.21)
6.64*
Triglycerides (mg/dL)
147.42 (120.47–191.17)
157.75 (136.69–176.66)
178.49 (146.89–207.69)
34.67***
Urea (mg/dL)
5.88 (4.90–7.03)
4.67 (3.79–5.79)
6.54 (5.08–8.19)
80.34***
Uric acid (mg/dL)
8.14 (7.17–9.20)
6.59 (5.38–8.07)
7.56 (5.80–9.51)
22.96***
Differences between moulting stages were tested using Kruskall-Wallis ANOVA (χ22). ns: non-significant; *p < 0.05; ***p < 0.001.
Table
2.
Results from Principal Component Analysis (using Varimax rotation) of plasma chemistry parameters (log-transformed) of adult Black-necked Grebes.
Parameter
PC1
PC2
PC3
PC4
Albumin
0.494
0.726
−0.210
−0.015
Alkaline phosphatase
0.070
0.094
−0.373
−0.239
β-hydroxy-butyrate
0.345
0.013
−0.097
0.463
Calcium
0.034
0.680
0.028
0.020
Cholesterol
0.848
0.226
−0.100
−0.043
Creatine kinase
0.244
−0.046
0.673
0.094
Glucose
0.388
0.124
−0.473
0.017
Lactate dehydrogenase
−0.026
−0.048
0.816
−0.114
Magnesium
−0.339
0.473
0.523
−0.298
Phosphorus
−0.125
0.200
0.350
0.609
Potassium
0.085
−0.101
0.091
0.642
Sodium
0.203
0.414
−0.094
−0.035
Total proteins
0.007
0.809
−0.052
−0.035
Triglycerides
0.718
0.184
−0.019
−0.052
Urea
0.550
−0.084
0.385
0.042
Uric acid
0.277
0.087
0.187
−0.603
% variance explained
14.8
13.9
13.4
9.6
Values for every parameter show loadings of the first four principal component (PC) axes. Values in bold are significant according to the brocken-stick criterion (Peres-Neto et al., 2003).
Table
3.
Generalised linear mixed models (GLMMs) of PC scores of adult Black-necked Grebes in relation to moulting stage (unmoulted, moulting, and moulted), date of capture and body condition (log-transformed scaled body mass).
Source of variation
F
Num df
Den df
P
PC1 scores
Moulting stage
8.12
2
660.7
< 0.001
Body condition
2.88
1
660.7
0.090
Date
268.38
1
660.5
< 0.001
PC2 scores
Moulting stage
6.79
2
649.8
0.001
Body condition
0.70
1
651.4
0.403
Date
1.18
1
658.8
0.279
PC3 scores
Moulting stage
2.76
2
660.8
0.064
Body condition
0.32
1
660.8
0.572
Date
31.63
1
660.5
< 0.001
The scores resulted from a Principal Component Analysis of plasma chemistry parameters. Numerator (Num) and Denominator (Den) of the df for the F values are given.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds