Abreu, C., Rocha-Pereira, N., Sarmento, A., Magro, F., 2015. Nocardia infections among immunomodulated inflammatory bowel disease patients: a review. World J. Gastroenterol. 21, 6491–6498.
|
Alberdi, A., Aizpurua, O., Bohmann, K., Zepeda-Mendoza, M.L., Gilbert, M.T.P., 2016. Do vertebrate gut metagenomes confer rapid ecological adaptation? Trends Ecol. Evol. 31, 689–699.
|
Alm, E.W., Daniels-Witt, Q.R., Learman, D.R., Ryu, H., Jordan, D.W., Gehring, T.M., et al., 2018. Potential for gulls to transport bacteria from human waste sites to beaches. Sci. Total Environ. 615, 123–130.
|
Amano, T., 2009. Conserving bird species in Japanese farmland: past achievements and future challenges. Biol. Conserv. 142, 1913–1921.
|
Amato, K.R., Leigh, S.R., Kent, A., Mackie, R.I., Yeoman, C.J., Stumpf, R.M., et al., 2015. The gut microbiota appears to compensate for seasonal diet variation in the wild black howler monkey ( Alouatta pigra). Microbial Ecol. 69, 434–443.
|
Bacciarini, L., Posthaus, H., Pagan, O., Miserez, R., 1999. Nocardia nova causing pulmonary nocardiosis of black crakes ( Limnocorax flavirostra). Vet. Pathol. 36, 345–347.
|
Bajaj, J.S., Ridlon, J.M., Hylemon, P.B., Thacker, L.R., Heuman, D.M., Smith, S., et al., 2012. Linkage of gut microbiome with cognition in hepatic encephalopathy. Am. J. Physiol.-Gastr. L. 302, G168-G175.
|
Baniel, A., Amato, K.R., Beehner, J.C., Bergman, T.J., Mercer, A., Perlman, R.F., et al., 2021. Seasonal shifts in the gut microbiome indicate plastic responses to diet in wild geladas. Microbiome 9, 1–20.
|
Barnes, E.M., 1972. The avian intestinal flora with particular reference to the possible ecological significance of the cecal anaerobic bacteria. Am. J. Clin. Nutr. 25, 1475–1479.
|
|
Barzen, J., 2012. Studying and understanding wetland dynamics. In: Li, F., Liu, G., Wu, J., Zeng, N., Harris, J., Jin, J. (Eds.), Ecological Study of Wetlands and Waterbirds at Poyang Lake. Popular Science Press, Beijing, pp. 256-266.
|
BirdLife International, 2022. Species factsheet: Leucogeranus leucogeranus. . (Accessed 29 November 2022).
|
Bolyen, E., Rideout, J.R., Dillon, M.R., Bokulich, N.A., Abnet, C.C., Al-Ghalith, G.A., et al., 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857.
|
Burrell, P., O'sullivan, C., Song, H., Clarke, W., Blackall, L., 2004. Identification, detection, and spatial resolution of Clostridium populations responsible for cellulose degradation in a methanogenic landfill leachate bioreactor. Appl. Environ. Microbiol. 70, 2414–2419.
|
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583.
|
Cao, L., Zhang, Y., Barter, M., Lei, G., 2010. Anatidae in eastern China during the non-breeding season: Geographical distributions and protection status. Biol. Conserv. 143, 650–659.
|
Chen, Y., Yang, F., Lu, H., Wang, B., Chen, Y., Lei, D., et al., 2011. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 54, 562–572.
|
|
China National Food Safety Standard, 2003. Determination of Crude Fiber in Vegetable Foods (GB/T 5009.10-2003). Ministry of Health of the People's Republic of China and Standardization Administration of the People's Republic of China, Beijing.
|
|
China National Food Safety Standard, 2010. Determination of Protein in Foods (GB 5009.5-2010). Ministry of Health of the People's Republic of China, Beijing.
|
|
China National Food Safety Standard, 2016a. Determination of Fat in Foods (GB 5009.6-2016). National Health and Family Planning Commission of the People's Republic of China and China Food and Drug Administration, Beijing.
|
|
China National Food Safety Standard, 2016b. Determination of Starch in Foods (GB 5009.9-2016). National Health and Family Planning Commission of the People's Republic of China and China Food and Drug Administration, Beijing.
|
|
China National Forestry Industry Standards, 1999. Determination of Crude Ash Content in Forest Plant and Forest Floor (LY/T 1268-1999). Chinese Academy of Forestry, Beijing.
|
Choi, C.-Y., Takekawa, J.Y., Xiong, Y., Ying, L., Wikelski, M., Heine, G., et al., 2016. Tracking domestic ducks: A novel approach for documenting poultry market chains in the context of avian influenza transmission. J. Integr. Agric. 15, 1584–1594.
|
Claesson, M.J., Jeffery, I.B., Conde, S., Power, S.E., O'connor, E.M., Cusack, S., et al., 2012. Gut microbiota composition correlates with diet and health in the elderly. Nature 488, 178–184.
|
Clarke, S.F., Murphy, E.F., O'Sullivan, O., Lucey, A.J., Humphreys, M., Hogan, A., et al., 2014. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 63, 1913–1920.
|
Czech, H.A., Parsons, K.C., 2002. Agricultural wetlands and waterbirds: a review. Waterbirds 25, 56–65.
|
Dai, X., Wan, R., Yang, G., 2015. Non-stationary water-level fluctuation in China's Poyang Lake and its interactions with Yangtze River. J. Geogr. Sci. 25, 274–288.
|
|
Darimont, C.T., Reimchen, T.E., Bryan, H.M., Paquet, P.C., 2008. Faecal-centric approaches to wildlife ecology and conservation: methods, data and ethics. Wildlife Biol. Pract. 4, 73–87.
|
David, L.A., Maurice, C.F., Carmody, R.N., Gootenberg, D.B., Button, J.E., Wolfe, B.E., et al., 2014. Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–563.
|
Davidson, N.C., 2014. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 65, 934–941.
|
Davidson, G.L., Wiley, N., Cooke, A.C., Johnson, C.N., Fouhy, F., Reichert, M.S., et al., 2020. Diet induces parallel changes to the gut microbiota and problem solving performance in a wild bird. Sci. Rep. 10, 20783.
|
|
Dhama, K., Mahendran, M., Tiwari, R., Singh, S.D., Kumar, D., Singh, S., et al., 2011. Tuberculosis in birds: insights into the Mycobacterium avium infections. Vet. Med. Int. 2011, 712369.
|
|
Di, M., Weng, X. Lu, P., Lu, Y., Dai, N., 2022. Dynamics of Grus leucogeranus in two farms in Poyang Lake during the winter of 2020-2021. Weland Sci. Manage. 18, 23–26.
|
Ezenwa, V.O., Gerardo, N.M., Inouye, D.W., Medina, M., Xavier, J.B., 2012. Animal behavior and the microbiome. Science 338, 198–199.
|
Fava, F., Gitau, R., Griffin, B.A., Gibson, G., Tuohy, K., Lovegrove, J., 2013. The type and quantity of dietary fat and carbohydrate alter faecal microbiome and short-chain fatty acid excretion in a metabolic syndrome 'at-risk'population. Int. J. Obes. 37, 216–223.
|
Feng, L., Hu, C., Chen, X., Cai, X., Tian, L., Gan, W., 2012. Assessment of inundation changes of Poyang Lake using MODIS observations between 2000 and 2010. Remote Sens. Environ. 121, 80–92.
|
Fontana, A., Manchia, M., Panebianco, C., Paribello, P., Arzedi, C., Cossu, E., et al. 2020. Exploring the role of gut microbiota in major depressive disorder and in treatment resistance to antidepressants. Biomedicines 8, 311.
|
Fox, A.D., Abraham, K.F., 2017. Why geese benefit from the transition from natural vegetation to agriculture. Ambio 46, 188–197.
|
Fox, A.D., Madsen, J., 2017. Threatened species to super-abundance: The unexpected international implications of successful goose conservation. Ambio 46, 179–187.
|
Fox, A.D., Elmberg, J., Tombre, I.M., Hessel, R., 2017. Agriculture and herbivorous waterfowl: A review of the scientific basis for improved management. Biol. Rev. 92, 854–877.
|
Franc, K., Krecek, R., Häsler, B., Arenas-Gamboa, A., 2018. Brucellosis remains a neglected disease in the developing world: a call for interdisciplinary action. BMC Publ. Health 18, 125.
|
Fu, R., Xiang, X., Dong, Y., Cheng, L., Zhou, L., 2020. Comparing the intestinal bacterial communies of sympatric wintering Hooded Crane ( Grus monacha) and Domestic Goose ( Anser anser domesticus). Avian Res. 11, 13.
|
Fuirst, M., Veit, R.R., Hahn, M., Dheilly, N., Thorne, L.H. 2018., Effects of urbanization on the foraging ecology and microbiota of the generalist seabird Larus argentatus. PLoS One 13, e0209200.
|
|
Ganz, H.H., Doroud, L., Firl, A.J., Hird, S.M., Eisen, J.A., Boyce, W.M., 2017. Community-level differences in the microbiome of healthy wild mallards and those infected by influenza A viruses. MSystems 2, e00188-00116.
|
Gao, B., Zhong, M., Shen, Q., Wu, Y., Cao, M., Ju, S., et al., 2020. Gut microbiota in early pregnancy among women with Hyperglycaemia vs. Normal blood glucose. BMC Pregnancy Childbirth 20, 284.
|
Gevers, D., Kugathasan, S., Denson, L.A., Vázquez-Baeza, Y., Van Treuren, W., Ren, B., et al., 2014. The treatment-naive microbiome in new-onset Crohn's disease. Cell Host Microbe 15, 382–392.
|
Gibbs, P.S., Kasa, R., Newbrey, J.L., Petermann, S.R., Wooley, R.E., Vinson, H.M., et al., 2007. Identification, antimicrobial resistance profiles, and virulence of members from the family Enterobacteriaceae from the feces of yellow-headed blackbirds ( Xanthocephalus xanthocephalus) in North Dakota. Avian Dis. 51, 649–655.
|
Gilbert, S.F., Bosch, T.C., Ledón-Rettig, C., 2015. Eco-Evo-Devo: developmental symbiosis and developmental plasticity as evolutionary agents. Nat. Rev. Genet. 16, 611–622.
|
Góngora, E., Elliott, K.H., Whyte, L., 2021. Gut microbiome is affected by inter-sexual and inter-seasonal variation in diet for thick-billed murres ( Uria lomvia). Sci. Rep. 11, 1–12.
|
Grond, K., Ryu, H., Baker, A.J., Santo Domingo, J.W., Buehler, D.M., 2014. Gastro-intestinal microbiota of two migratory shorebird species during spring migration staging in Delaware Bay, USA. J. Ornithol. 155, 969–977.
|
Grond, K., Sandercock, B.K., Jumpponen, A., Zeglin, L.H., 2018. The avian gut microbiota: community, physiology and function in wild birds. J. Avian Biol. 49, e01788.
|
Hall, A.B., Tolonen, A.C., Xavier, R.J., 2017. Human genetic variation and the gut microbiome in disease. Nat. Rev. Genet. 18, 690–699.
|
Harris, J., Mirande, C., 2013. A global overview of cranes: status, threats and conservation priorities. Chinese Birds 4, 189–209.
|
|
He, W., Shao, M., Zhi, Y., Dai, N., Lu, P., Tu, X., 2019. Waterbird diversity in three reclamation regions of Poyang Lake. Chinese J. Ecol. 38, 2765–2771.
|
Hemminger, K., König, H., Månsson, J., Bellingrath-Kimura, S.D., Nilsson, L., 2022. Winners and losers of land use change: A systematic review of interactions between the world's crane species (Gruidae) and the agricultural sector. Ecol. Evol. 12, e8719.
|
Herlemann, D., Labrenz, M., Jürgens, K., Bertilsson, S., Waniek, J.J., Andersson, A.F., 2011. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 5, 1571–1579.
|
Hicks, A.L., Lee, K.J., Couto-Rodriguez, M., Patel, J., Sinha, R., Guo, C., et al., 2018. Gut microbiomes of wild great apes fluctuate seasonally in response to diet. Nat. Commun. 9, 1–18.
|
|
Hird, S.M., Sánchez, C., Carstens, B.C., Brumfield, R.T., 2015. Comparative gut microbiota of 59 neotropical bird species. Front. Microbiol. 6, 1403.
|
Hooper, L.V., Littman, D.R., Macpherson, A.J., 2012. Interactions between the microbiota and the immune system. Science 336, 1268–1273.
|
Hou, J., Wang, Y., Jin, B., Wang, L., Wang, W., 2019. Food composition of Siberian Cranes in agricultural fields in the Poyang Lake, China. Chin. J. Zool. 54, 15–21.
|
Hou, J., Liu, Y., Fraser, J.D., Li, L., Zhao, B., Lan, Z., et al., 2020. Drivers of a habitat shift by critically endangered Siberian cranes: Evidence from long-term data. Ecol. Evol. 10, 11055–11068.
|
Hou, J., Li, L., Wang, Y., Wang, W., Zhan, H., Dai, N., et al., 2021. Influences of submerged plant collapse on diet composition, breadth, and overlap among four crane species at Poyang Lake, China. Front. Zool. 18, 1–17.
|
|
Hu, Z., Lin, Y., 2019. Analysis of evolution process and driving factors for aquatic vegetations of Poyang Lake in 30 years. Resour. Environ. Yangtze Basin 28, 1947–1955.
|
|
Jia, Y., 2013. Impact of Water Level Fluctuation on Siberian Crane and Other Wintering Waterbirds in Poyang Lake. Doctoral Thesis. Beijing Forestry University, Beijing.
|
Jia, Y., Jiao, S., Zhang, Y., Zhou, Y., Lei, G., Liu, G., 2013. Diet shift and its impact on foraging behavior of Siberian crane ( Grus leucogeranus) in Poyang Lake. PLoS One 8, e65843.
|
Kim, H.I., Hong, S.H., Lee, S.Y., Ku, J.M., Kim, M.J., Ko, S.-G., 2021. Gardenia jasminoides ameliorates antibiotic-associated aggravation of DNCB-induced atopic dermatitis by restoring the intestinal microbiome profile. Nutrients 13, 1349.
|
Knutie, S.A., Chaves, J.A., Gotanda, K.M., 2019. Human activity can influence the gut microbiota of Darwin's finches in the Galapagos Islands. Mol. Ecol. 28, 2441–2450.
|
Kobayashi, T., Osaki, T., Oikawa, S., 2015. Use of T-RFLP and seven restriction enzymes to compare the faecal microbiota of obese and lean Japanese healthy men. Benef. Microbes 6, 735–745.
|
Le Bastard, Q., Al-Ghalith, G., Grégoire, M., Chapelet, G., Javaudin, F., Dailly, E., et al., 2018. Systematic review: human gut dysbiosis induced by non-antibiotic prescription medications. Aliment Pharmacol. Therapeut. 47, 332–345.
|
Le Chatelier, E., Nielsen, T., Qin, J., Prifti, E., Hildebrand, F., Falony, G., et al., 2013. Richness of human gut microbiome correlates with metabolic markers. Nature 500, 541–546.
|
Lewis, W.B., Moore, F.R., Wang, S. 2017. Changes in gut microbiota of migratory passerines during stopover after crossing an ecological barrier. Auk 134, 137–145.
|
Ley, R.E., Hamady, M., Lozupone, C., Turnbaugh, P.J., Ramey, R.R., Bircher, J.S., et al., 2008. Evolution of mammals and their gut microbes. Science 320, 1647–1651.
|
Li, Y., Qian, F., Silbernagel, J., Larson, H., 2019. Community structure, abundance variation and population trends of waterbirds in relation to water level fluctuation in Poyang Lake. J. Great Lake. Res. 45, 976–985.
|
Li, M., Wang, B., Zhang, M., Rantalainen, M., Wang, S., Zhou, H., et al., 2008. Symbiotic gut microbes modulate human metabolic phenotypes. P. Natl. Acad. Sci. 105, 2117–2122.
|
|
Li, Y., Huang, J., Li, F., Yulin, O., 2013. Diurnal time budget and behavior rhythm of wintering tundra swan after higher water level in Poyang Lake. Sichuan J. Zool. 32, 498–503.
|
Li, G., Yin, B., Li, J., Wang, J., Wei, W., Bolnick, D.I., et al., 2020. Host-microbiota interaction helps to explain the bottom-up effects of climate change on a small rodent species. ISME J. 14, 1795–1808.
|
Li, G., Li, J., Kohl, K.D., Yin, B., Wei, W., Wan, X., et al., 2019. Dietary shifts influenced by livestock grazing shape the gut microbiota composition and co-occurrence networks in a local rodent species. J. Anim. Ecol. 88, 302–314.
|
Li, C., Liu, Y., Gong, M., Zheng, C., Zhang, C., Li, H., et al., 2021. Diet-induced microbiome shifts of sympatric overwintering birds. Appl. Microbiol. Biotechnol. 105, 5993–6005.
|
Li, Y., Zhong, Y., Shao, R., Yan, C., Jin, J., Shan, J., et al., 2020. Modified hydrological regime from the three gorges dam increases the risk of food shortages for wintering waterbirds in Poyang Lake. Global Ecol. Conserv. 24, e01286.
|
Long, P., Choi, G., Silberman, M., 1983. Nocardiosis in two Pesquet's parrots ( Psittrichas fulgidus). Avian Dis. 855–859.
|
Lu, Z., Li, S., Wang, M., Wang, C., Meng, D., Liu, J., 2022. Comparative analysis of the gut microbiota of three sympatric terrestrial wild bird species overwintering in farmland habitats. Front. Microbiol. 13, 905668.
|
Macke, E., Tasiemski, A., Massol, F., Callens, M., Decaestecker, E., 2017. Life history and eco-evolutionary dynamics in light of the gut microbiota. Oikos 126, 508–531.
|
Mahtab, N., Zhou, L., Zhang, F., Wang, W., 2021. Seasonal variations in the gut fungal communities of Hooded Crane ( Grus monacha) at wintering and stopover sites in China. Animals 11, 941.
|
Manichanh, C., Rigottier-Gois, L., Bonnaud, E., Gloux, K., Pelletier, E., Frangeul, L., et al., 2006. Reduced diversity of faecal microbiota in Crohn's disease revealed by a metagenomic approach. Gut 55, 205–211.
|
Margiotta, E., Caldiroli, L., Callegari, M.L., Miragoli, F., Zanoni, F., Armelloni, S., et al., 2021. Association of sarcopenia and gut microbiota composition in older patients with advanced chronic kidney disease, investigation of the interactions with uremic toxins, inflammation and oxidative stress. Toxins 13, 472.
|
McMurdie, P.J., Holmes, S., 2013. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS one 8, e61217.
|
Medawar, E., Haange, S.-B., Rolle-Kampczyk, U., Engelmann, B., Dietrich, A., Thieleking, R., et al., 2021. Gut microbiota link dietary fiber intake and short-chain fatty acid metabolism with eating behavior. Transl. Psychiatry 11, 1–11.
|
Meng, W., He, M., Hu, B., Mo, X., Li, H., Liu, B., et al., 2017. Status of wetlands in China: A review of extent, degradation, issues and recommendations for improvement. Ocean Coast Manag. 146, 50–59.
|
Meng, S., Chen, B., Yang, J., Wang, J., Zhu, D., Meng, Q., et al., 2018. Study of microbiomes in aseptically collected samples of human breast tissue using needle biopsy and the potential role of in situ tissue microbiomes for promoting malignancy. Front. Oncol. 8, 318.
|
Michel, A.J., Ward, L.M., Goffredi, S.K., Dawson, K.S., Baldassarre, D.T., Brenner, A., et al., 2018. The gut of the finch: uniqueness of the gut microbiome of the Galápagos vampire finch. Microbiome 6, 1–14.
|
Moono, P, Foster, N.F., Hampson, D.J., Knight, D.R., Bloomfield, L.E., Riley, T.V., 2016. Clostridium difficile infection in production animals and avian species: a review. Foodb. Pathog. Dis. 13, 647–655.
|
Muegge, B.D., Kuczynski, J., Knights, D., Clemente, J.C., González, A., Fontana, L., et al., 2011. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 332, 970–974.
|
Murray, M.H., Lankau, E.W., Kidd, A.D., Welch, C.N., Ellison, T., Adams, H.C., et al., 2020. Gut microbiome shifts with urbanization and potentially facilitates a zoonotic pathogen in a wading bird. PLoS One 15, e0220926.
|
Muzaffar, S.B., Takekawa, J.Y., Prosser, D.J., Newman, S.H., Xiao, X., 2010. Rice production systems and avian influenza: interactions between mixed-farming systems, poultry and wild birds. Waterbirds 33, 219–230.
|
Nagy-Szakal, D., Hollister, E.B., Luna, R.A., Szigeti, R., Tatevian, N., Smith, C.W., et al., 2013. Cellulose supplementation early in life ameliorates colitis in adult mice. PloS one 8, e56685.
|
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., O'hara, R., et al., 2013. vegan: Community Ecology Package. .
|
Pan, L., Han, P., Ma, S., Peng, R., Wang, C., Kong, W., et al., 2020. Abnormal metabolism of gut microbiota reveals the possible molecular mechanism of nephropathy induced by hyperuricemia. Acta Pharm. Sin. B 10, 249–261.
|
Pfeiler, E.A., Klaenhammer, T.R., 2007. The genomics of lactic acid bacteria. Trends Microbiol. 15, 546–553.
|
Phillips, G., Willby, N., Moss, B., 2016. Submerged macrophyte decline in shallow lakes: what have we learnt in the last forty years? Aquat. Bot. 135, 37–45.
|
Prosser, D.J., Palm, E.C., Takekawa, J.Y., Zhao, D., Xiao, X., Li, P., et al., 2016. Movement analysis of free-grazing domestic ducks in Poyang Lake, China: a disease connection. Int. J. Geogr. Inf. Sci. 30, 869–880.
|
|
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al., 2013. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 41, D590-D596.
|
|
Salamon, D., Sroka-Oleksiak, A., Kapusta, P., Szopa, M., Mrozińska, S., Ludwig-Słomczyńska, A.H., et al., 2018. Characteristics of gut microbiota in adult patients with type 1 and type 2 diabetes based on next-generation sequencing of the 16S rRNA gene fragment. Pol. Arch. Intern. Med. 128, 336–343.
|
Scanu, A.M., Bull, T.J., Cannas, S., Sanderson, J.D., Sechi, L.A., Dettori, G., et al., 2007. Mycobacterium avium subspecies paratuberculosis infection in cases of irritable bowel syndrome and comparison with Crohn's Disease and Johne's Disease: common neural and immune pathogenicities. J. Clin. Microbiol. 45, 3883–3890.
|
|
Shao, M., Gong, H., Dai, N., Zhi, Y., Xu, N., Lu, P., 2018. Study on time budgets and behavioral rhythm of wintering Siberian cranes in a lotus pond reclamation area in Poyang Lake. Acta Ecol. Sin. 38, 5206–5212.
|
Tang, X., Zhang, L., Fan, C., Wang, L., Fu, H., Ren, S.E., et al., 2021. Dietary fiber influences bacterial community assembly processes in the gut microbiota of Durco×Bamei Crossbred Pig. Front. Microbiol. 12, 688554.
|
Teyssier, A., Matthysen, E., Hudin, N.S., De Neve, L., White, J., Lens, L., 2020. Diet contributes to urban-induced alterations in gut microbiota: experimental evidence from a wild passerine. Proc. R. Soc. B 287, 20192182.
|
Van Dyke, M., McCarthy, A., 2002. Molecular biological detection and characterization of Clostridium populations in municipal landfill sites. Appl. Environ. Microbiol. 68, 2049–2053.
|
Van Immerseel, F., Rood, J.I., Moore, R.J., Titball, R.W., 2009. Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens. Trends Microbiol. 17, 32–36.
|
Van Veelen, H.P.J., Salles, J.F., Tieleman, B.I., 2017. Multi-level comparisons of cloacal, skin, feather and nest-associated microbiota suggest considerable influence of horizontal acquisition on the microbiota assembly of sympatric woodlarks and skylarks. Micobiome 5, 156.
|
Videvall, E., Song, S.J., Bensch, H.M., Strandh, M., Engelbrecht, A., Serfontein, N., et al., 2019. Major shifts in gut microbiota during development and its relationship to growth in ostriches. Mol. Ecol. 28, 2653–2667.
|
|
Waite, D.W., Taylor, M.W., 2014. Characterizing the avian gut microbiota: membership, driving influences, and potential function. Front. Microbiol. 5, 223.
|
Wang, Q., Garrity, G.M., Tiedje, J.M., Cole, J.R., 2007. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267.
|
Wang, W., Fraser, J.D., Chen, J., 2017. Wintering waterbirds in the middle and lower Yangtze River floodplain: changes in abundance and distribution. Bird Conserv. Int. 27, 167–186.
|
Wang, W., Wang, L., Hou, J., 2019. Man-made habitats have become important foraging areas of Siberian cranes. Chinese J. Wildlife 40, 133–137.
|
Wang, Q., Zhao, L., Bin, Y., Wang, L., Wang, L., Zhang, K., et al., 2022. Blood bacterial 16S rRNA gene alterations in women with polycystic ovary syndrome. Front. Endocrinol. 13, 814520.
|
West, A.G., Waite, D.W., Deines, P., Bourne, D.G., Digby, A., McKenzie, V.J., et al., 2019. The microbiome in threatened species conservation. Biol. Conserv. 229, 85–98.
|
|
Wickham, H., 2009. ggplot2: Elegant Graphics for Data Analysis. Springer, New York.
|
Wooley R, Gibbs P, Brown T, Maurer J., 2000. Chicken embryo lethality assay for determining the virulence of avian Escherichia coli isolates. Avian Dis. 44, 318–324.
|
|
Wu, J., Li, F., Burnham, J., 2013. Numerical distribution of Siberian cranes and their relation to food and water depth in Sha Hu at Poyang Lake, China. Wetland Sci. 11, 305–312.
|
Wu, Q., Wang, X., Ding, Y., Hu, Y., Nie, Y., Wei, W., et al., 2017. Seasonal variation in nutrient utilization shapes gut microbiome structure and function in wild giant pandas. Proc. R. Soc. A B 284, 20170955.
|
Xiang, X., Zhang, F., Fu, R., Yan, S., Zhou, L., 2019. Significant differences in bacterial and potentially pathogenic communities between sympatric hooded crane and greater white-fronted goose. Front. Microbiol. 10, 163.
|
Xing, Y., Wang, D., 2016. Research progress on surveillance related to avian influenza activity in the Poyang Lake Region of China. Chin. J. Zoonoses 32, 388–391.
|
Yang, Z., Zhou L., 2021. Is intestinal bacterial diversity enhanced by trans-species spread in the mixed-species flock of Hooded Crane ( Grus monacha) and Bean Goose ( Anser fabalis) wintering in the lower and middle Yangtze River floodplain? Animals 11, 233.
|
Zhang, Y., Jeppesen, E., Liu, X., Qin, B., Shi, K., Zhou, Y., et al., 2017. Global loss of aquatic vegetation in lakes. Earth Sci. Rev. 173, 259–265.
|
Zhang, H., Chang, Y., Zheng, Q., Zhang, R., Hu, C., Jia, W., 2019. Altered intestinal microbiota associated with colorectal cancer. Front. Med. 13, 461–470.
|
Zhang, F., Xiang, X., Dong, Y., Yan, S., Song, Y., Zhou, L., 2020. Significant differences in the gut bacterial communities of Hooded Crane ( Grus monacha) in different seasons at a stopover site on the flyway. Animals 10, 701.
|
|
Zhong, Y., 2020. Diversity and Habitat Selection of Birds in Poyang Lake Rice Fields. Master's Thesis. . Jiangxi Normal University, Nanchang.
|
Zhu, Y., Li, Y., Yang, H., He, K., Tang, K., 2021. Establishment of gut microbiome during early life and its relationship with growth in endangered crested ibis ( Nipponia nippon). Front. Microbiol. 12, 723682.
|

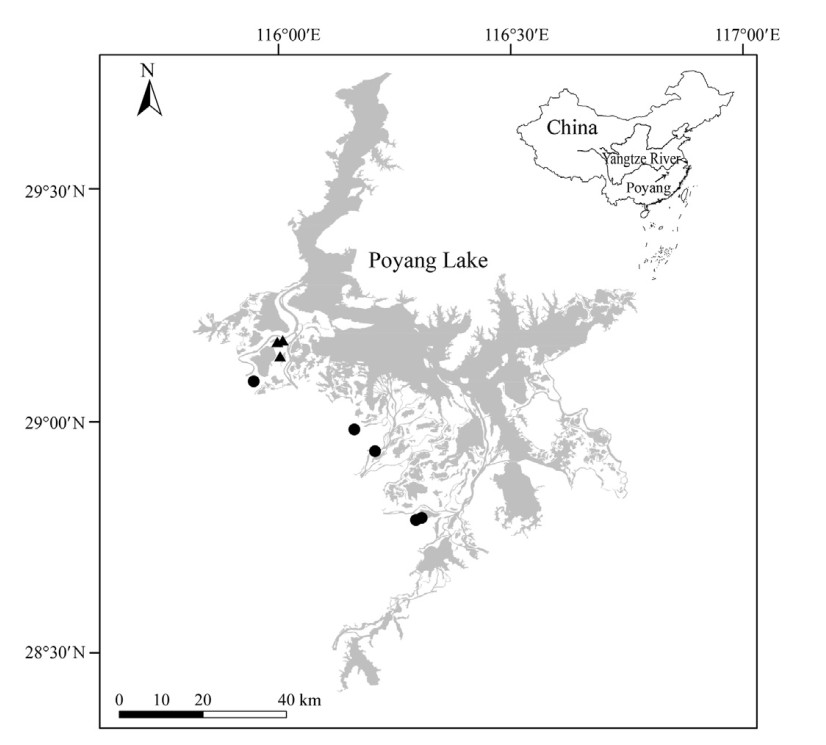
 DownLoad:
DownLoad:
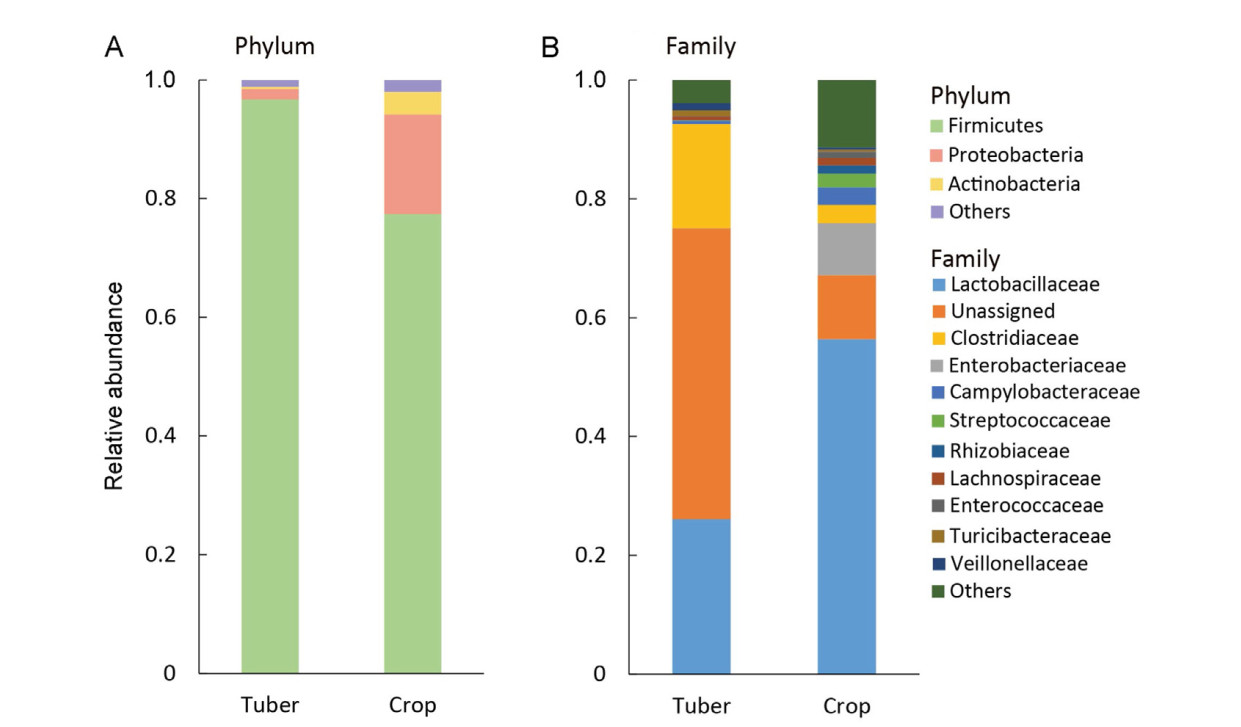
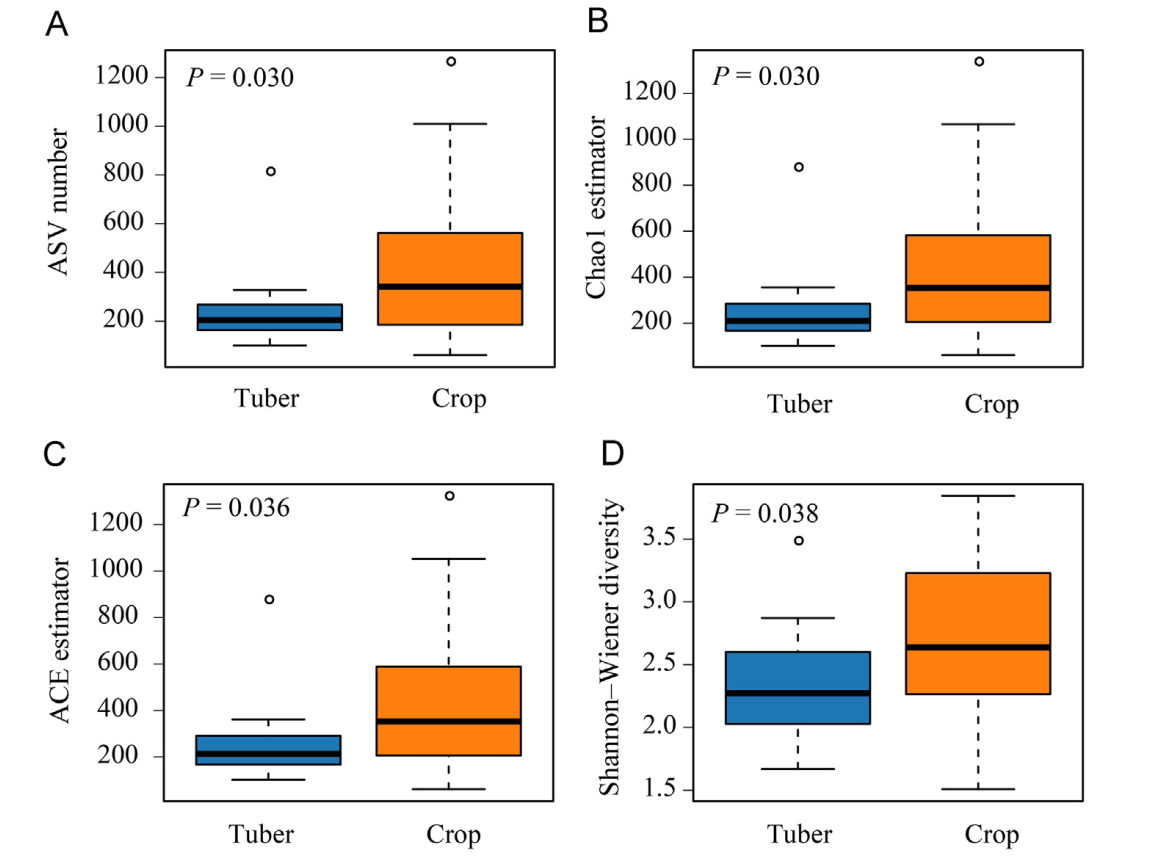
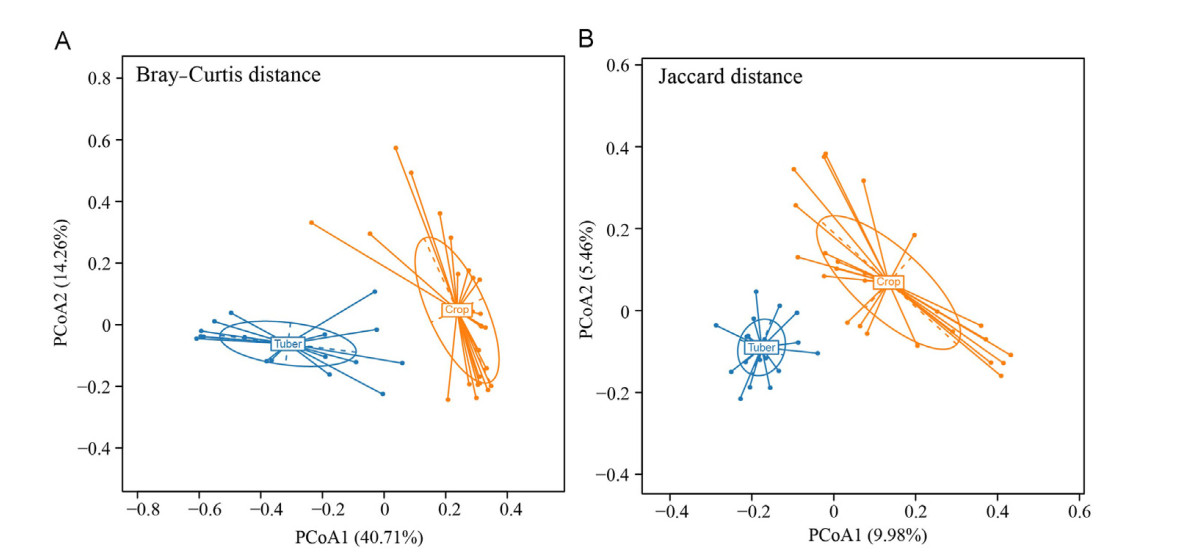





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds