Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China
b.
College of Life Science, University of Chinese Academy of Sciences, Beijing, 100049, China
c.
Center for Macroecology, Evolution and Climate, Globe Institute, University of Copenhagen, DK-2100, Copenhagen, Denmark
d.
Center for Excellence in Animal Evolution and Genetics, Chinese Academy of Sciences, Kunming, 650223, China
Funds: This study was financially supported by the National Key Research and Development Program of China (Grant No. 2022YFC2601601), the National Natural Science Foundation of China (Grant No. 31727901) and the China Scholarship Council (No. 2019–13045 to H.X.)
Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China. E-mail address: leifm@ioz.ac.cn (F. Lei)
With the continued development of tracking technology and increasing interest in animal movement, our understanding of migration behavior has become more comprehensive. However, there are still many species that have not been well studied, particularly sea birds. Here, we present the first year-round Global Positioning System (GPS) tracking data of the Black-tailed Gull (Larus crassirostris) at the population level. We used solar-powered GPS-Global System for Mobile communication (GSM) loggers to successfully track 30 individuals breeding at Xingrentuo Islet, Liaoning Province, China, for 1–3 years. Except for one individual who roamed in the far north of the Yellow Sea during non-breeding period, all others did a directed southward migration. Migration routes and wintering sites differed among migrating gulls and between years for the birds tracked for two or more years. Additionally, during wintering, the migrating gulls were more likely to travel over a large body of water and shift sites, and some trajectories were quite complex, which was probably closely related to what we observed in the field about their boat-chasing behavior. Compared to wintering movements, the post-breeding movements ranged over a smaller area. Specifically, almost all of them had a long post-breeding period near the breeding islet (≥120 days, <220 km from the breeding islet), and 80% of the gulls who were tracked more than one year had at least one faithful post-breeding site. Compared to the post-breeding period, only approximately half of the migrating gulls had a pre-breeding period that was shorter (3–20 days) and closer to the breeding islet (≤80 km). Migration distance varied among migrating gulls (range 209–2405 km) and the gulls moved least distance during post-breeding period. Furthermore, we found that the southward movement of the migrating gulls occurred when the temperature near the breeding islet dropped; specifically, the gulls directly migrated southward away from the post-breeding site. Our results suggest that the Black-tailed Gull has a long post-breeding period but a short pre-breeding period near the breeding islet and high diversity of their migrating patterns (in especial migration routes and wintering sites).
From an evolutionary standpoint, bare-throated spurfowls (Pternistis spp.) in Africa were probably originally birds of acacia habitat, first split by the spread of woodlands and forests (Hall, 1963). This split created the proto-yellow-throated (proto-leucosceptus) group in East Africa and the proto-red-throated (proto-afer/swainsonii) group in the southern parts of Africa (Hall, 1963). At least four species developed: the Yellow-necked Spurfowl (P. leucosceptus; with yellow throat), Red-necked Spurfowl (P. afer; with scarlet to red throats), Grey-breasted Spurfowl (P. rufopictus; with orange throats) and Swainson's Spurfowl (P. swainsonii; with scarlet red throats). Recently Mandiwana-Neudani et al. (2019) have elevated a subspecies of the Red-necked Spurfowl, namely P. afer cranchii, to full species status and therefore recognise five bare-throated spurfowls. However, spurfowl taxonomy is currently the subject of revision, including the subspecies of P. afer (Crowe et al., 2022). In this paper, P. afer cranchii is used to represent the Red-necked Spurfowl. The remainder of the spurfowls in Africa, about 19 species, have feathered throats (Madge and McGowan, 2001).
Spurfowl are generally cryptic ground-living landfowl (Madge and McGowan, 2001). Their cryptic status may explain why their bare throats have never become the focal point of scientific endeavour (cf. Iverson and Karubian, 2017). However, from a bird's perspective, even small and localised ornamental features could have a vital signalling function, achieving the same purpose as the extravagant ornamentation used for courtship in birds such as peacocks (Iverson and Karubian, 2017). For example, the highly cryptic Crested Francolin (Dendroperdix sephaena) female has fine transverse bars on her back, which she erects to attract the male during courtship (van Niekerk, 2001). The Chinese Grouse (Tetrastes sewerzowi) male is dull without extravagant colours. Still, the males with more extensive and brighter red supra-orbital combs gain access to breeding females, although it is a small ornament (Yang et al., 2013). During the mating season, the red colouration of the beak, eye rings and legs in Red-legged Partridge (Alectoris rufa) is brighter due to pigments from dietary carotenoids, and there is evidence that males use these traits for signalling during courtship (Perez-Rodríguez et al., 2013).
Both sexes of the bare-throated spurfowls have similarly coloured bare throats, but males develop saturated throat colours. Otherwise, they are described as sexually monomorphic (Madge and McGowan, 2001). From an evolutionary perspective, it is intriguing, since both sexes have acquired bare throats, unlike other spurfowl with feathered throats, which possibly facilitate thermoregulation under severely hot conditions. The spurfowls probably use a network of arteries in the throat skin to cool down the brain by dissipating heat (cf. van Niekerk et al., 2022). However, unlike females, males also use these coloured throat patches for territorial advertisement displays (van Niekerk, 2017).
Male birds' acquisition of bright plumage to attract females has received considerable coverage since Charles Darwin published his famous theory on sexual selection 151 years ago (Petrie, 2021). It is generally accepted that females use male plumage colouration as a criterion in mate choice (Liu et al., 2009). However, less focus has been given to bare facial or neck patches as a signal used by females (Iverson and Karubian, 2017). Determining the purpose of colourful bare throats in spurfowls has not yet been considered, even though crowing territorial males boldly show these features: during courtship, they sit on elevated posts, turn their head upwards, inflate their colourful throats and crow loudly (van Niekerk, 1983, 2017; van Niekerk et al., 2009).
The effect of the presence of ectoparasites on ornaments is a novelty from a signalling point of view. Bare-throated spurfowls have the propensity to attract ectoparasites to their bare throats. Since the colour condition of throats may play a role in reproductive behaviour, it is essential to understand if ectoparasites affect this male-female interaction. Among Satin Bowerbirds (Ptilonorhynchus violaceus), a correlated infection model suggests that females favour males with lower levels of ectoparasite infestation to keep their resistance to internal infections low (Borgia and Collis, 1989). However, this study by Borgia and Collins does not reveal information on the optics of ectoparasites on skin ornaments that may play a negative role during male–female interaction.
The goals of this study are (1) to describe the function of the bare throats in the breeding biology of the bare-throated spurfowls; (2) to determine why females have colourful throats, seeing that this may not have any sexual significance; (3) to describe the association between different throat colours (and degree of saturation) and levels of ectoparasite infestation and (4) to determine whether the bare throat sizes of males result in sexual size dimorphism.
2.
Materials and methods
Since the colours had faded completely on museum skins, I could only conduct a colour assessment of spurfowl bare throats of suitable photographs (Kennedy, 2010). Yang et al. (2013) and Laitly et al. (2012) showed how useful citizen science photographs are for biodiversity research. My data were sourced from the online Macaulay Library in the Cornell Lab of Ornithology (ML) (https://www.macaulaylibrary.org/). The selected photographs were taken in national parks/game reserves, mostly from safari vehicles making camera height (1–1.5 m) and distance (10–20 m) consistent. The spurfowls were habituated to humans and were mostly photographed on or near the road in their natural habitats along fixed routes. These experienced photographers mainly used sophisticated SLR Canon or Nikon cameras; the lens focal lengths were 400–500 mm. Flashes did not fire off. ML checked each entry for originality and quality before entering. The quality ranking was from 1 to 5, with the criteria described by ML. The selected material was rated 4 to 5 with high resolution to allow zooming.
Variable light intensity and time of day can compromise the true colours of birds in photographs (Withers and Hosking, 1996). The following guidelines were applied when selecting photographs for throat colour assessment:
1. Only photographs with clearly visible throats were selected.
2. Swainson's Spurfowl males with saturated red pigmentation showed a thickish "double chin" lacking in the other species.
3. Richly saturated throats were distinguishable from poorly coloured ones where several birds occurred in the same photograph.
4. Saturation often corresponds with the rainy season.
5. Saturation invariably corresponded with saturated beak and leg colours in the Red-necked Spurfowl.
6. The bare orbital ring in all species matched the bare throat colour regarding pigment saturation.
7. Saturated and less saturated patches were found on the orbital skin of the same bird. This clearly shows that bright and vague colours can be distinguished in the same photograph.
8. Bleached throats (not saturated) corresponded with other run-down conditions such as washed-out or rugged leg colours and integument.
Unlike the other three species, Red-necked Spurfowl appears to shrink the size of their bare throats to less than 50% of the expected size. Peripheral throat feathers cover part of the shrunk throat skin during cold, windy conditions (JHvN personal observations). Therefore, these photographs were excluded from the analyses.
The correlated monthly rainfall and throat colour intensity was also investigated in relation to breeding (nests with eggs). Relating this to Swainson's Spurfowl distribution, rainfall data for the Kruger National Park (South Africa), Harare (Zimbabwe), Lusaka (Zambia) and Otjiwarongo (Namibia) was averaged (http://www.meteoblue.com). For the area inhabited by Red-necked Spurfowls, monthly rainfall data from Tanzania, Uganda and Kenya were averaged (http://www.meteoblue.com). The monthly rainfall data from Weather and Climate in Serengeti National Park (http://www.serenegti.com) applied to Yellow-necked and Grey-breasted Spurfowls.
The identification number of photographs (Macaulay Library) and the pigmentation scores (degree of saturation) are captured in Appendix Table S1, S2, S3, S4. The throat colours of each photo, covering four species, were divided into four categories/weighted scores depending on the intensity of colour pigmentation (Table 1). This approach was made for each month of the year to correlate throat saturation with monthly rainfall and monthly nests with eggs data from the Tanzanian Bird Atlas (http://tanzaniabirdatlas.net/start.htm) to the monthly degree of throat colour saturation of Red-necked, Yellow-necked and Grey-breasted Spurfowls. Liversage (1978) was consulted for nest data that would apply to Swainson's Spurfowl.
Table
1.
Weighted scores were assigned to colour pigment intensity of the bare throat of the bare-throated spurfowls (see Appendix Table S1, S2, S3, S4).
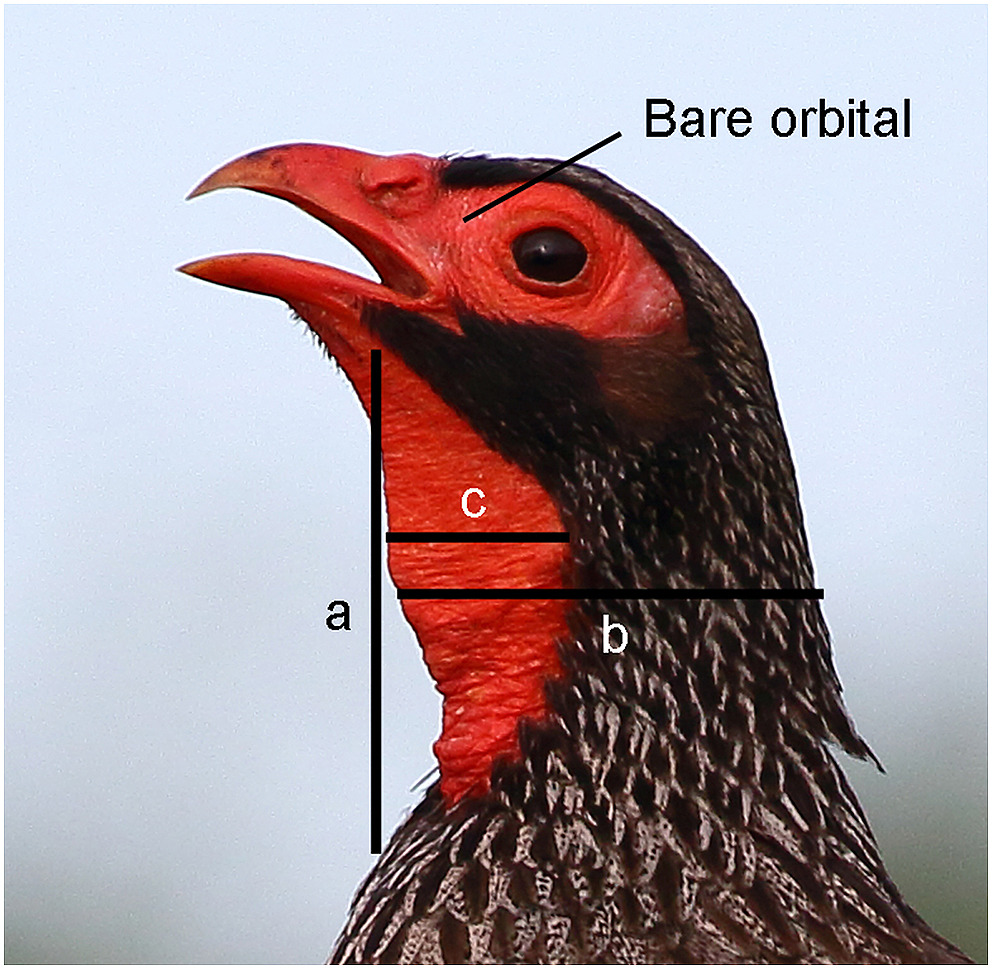
Besides colour assessment, measurements from photographs were also obtained to determine bare throat size differences among species for sexual size dimorphism. In the end, 96 suitable pictures of male and female throats were assessed representing all four species (Appendix Table S5). These photographs were all lateral views so that it was possible to measure the length of the throat (a), the width of the bare throat (c) and the width of the neck (b) with a vernier in mm on the computer screen (Fig. 1).
Figure
1.
Three measurements were taken from the lateral aspect of bare-throated spurfowl photographs to determine the relative surface area of the bare part (%) (Photo credit: Moishie Hersko).
Body size of males and females engaged in courtship displays from all suitable photographs were also measured in mm with vernier calipers (Table 2). In the same picture, two dimensions were selected: the height and width of the male and female bodies engaged in courtship behaviour. These data were obtained for bare-throated spurfowls and feather-throated spurfowls, so it was possible to compare the two data sets (Table 2). The data were converted and expressed as percentage differences.
Table
2.
Width and height of males and females of both bare-throated and feather-throated spurfowls converted to percentage differences between the two sub-groups of spurfowl (see materials and methods).
The basic ectoparasite assessment appears in Appendix Table S1, S2, S3, S4. Ectoparasites were recorded as present or absent on the bare throat of spurfowls. These scores allowed the determination of whether ectoparasites discriminated between colours (Table 3). This primary habitat determinant for the different bare-throated species revolved around tree canopy. The tree canopy density in various African localities was obtained from Hanan et al. (2020) and annual mean temperatures from Nwaerema et al. (2019).
Table
3.
Ectoparasite scores detected on the throats of the bare-throated spurfowls tabulated in four colour categories.
Colour saturation
1
2
3
4
Males
No. of specimens
With ectoparasites
No. of specimens
With ectoparasites
No. of specimens
With ectoparasites
No. of specimens
With ectoparasites
P. a. swainsonii
1
0
33
1
73
1
72
1
P. a. leucoscepus
10
6
54
16
71
13
29
8
P. s. ruficollis
1
0
23
3
25
3
20
0
P. a. cranchii
9
1
38
4
42
0
5
0
Totals
21
7
148
24
211
17
126
9
Percentages
33.33
16.21
8.05
7.14
Females
P. a. swainsonii
5
0
51
1
45
1
13
0
P. a. leucoscepus
20
3
38
10
17
5
5
1
P. s. ruficollis
6
0
19
3
9
0
4
1
P. a. cranchii
18
0
33
0
4
0
0
0
Totals
49
3
141
14
75
6
22
2
Percentages
6.12
9.92
8
9.09
Colour saturation: 1 = very bleached, 2 = bleached, 3 = saturated, 4 = highly saturated (see Table 1).
For the assessment of throat colours, each photograph received a score from 1 to 4. I converted this data into frequency tables, e.g., how many 1's, 2's, 3's and 4's fell in each month of the year (see Appendix Table S6). These frequencies were then converted to percentages using Excel. The 3% and 4% frequencies were regarded as saturated colours and added up to get one monthly value (see Appendix Table S7). The 1 and 2 frequencies were not bright. These monthly percentages of the saturated colours (3's and 4's added) were correlated with average monthly rainfall and egg-laying data using Pearson's correlation (Statistics Kingdom). All data sets were checked for normality using the Shapiro-Wilk test (alfa = 0.05) (Statistics Kingdom).
The bare throat size is a percentage of the total neck area (throat area = a × c and total neck area = a × b, see Fig. 1). I used two-way ANOVA (Statistics Kingdom) to determine the throat size differences between species. I compared the height and width of the sexes in the photographs for courtship displays and converted them to percentage differences. For height, the formula is [(male height − female height)/male height] × 100, and for width [(male width − female width)/male width] × 100 (Table 3).
3.
Results
3.1
Seasonal throat colour variation, breeding and rainfall
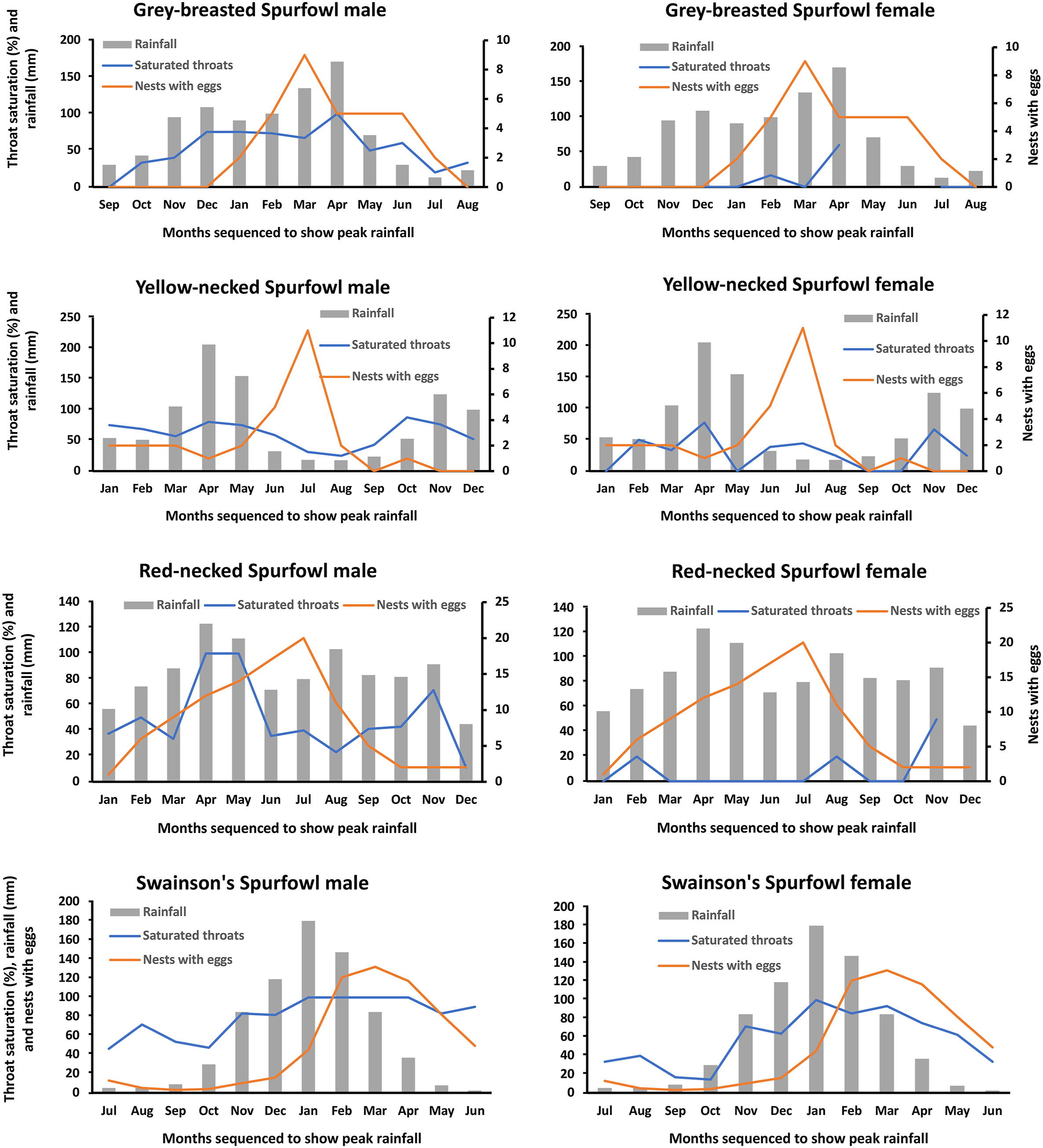
Different throat colour intensities of males correlated significantly with average monthly rainfall figures in the males of all four species and the female of Swainson's Spurfowl (Fig. 2; Table 4). In three species, i.e., Yellow-necked, Red-necked and Grey-breasted Spurfowls, throat saturation did not correlate with egg laying but occurred one or two months earlier. Among Swainson's Spurfowl males and females, saturation correlated with monthly egg laying. In three species, except Grey-breasted Spurfowl, females produced eggs immediately after the peak rainy months of the year. Grey-breasted Spurfowls lay eggs during the peak rainy months (Fig. 2). The saturated throat colours of the males are shown in Fig. 3, and examples of the bleached throat are in Fig. 4.
Figure
2.
Correlation between monthly rainfall and spurfowl throat saturation.
Table
4.
Correlation statistics between monthly throat colour saturation vs rainfall and monthly throat colour saturation vs. nest records of four bare-throated spurfowl species.
Figure
3.
Saturated throats of the bare-throated spurfowls. Left to right: Red-necked (P. afer cranchii; photo credit Michael Orter), Yellow-throated (P. leucosceptus; photo credit Peter Candido), Grey-breasted (P. rufopictus; photo credit Markus Lilijer) and Swainson's (P. swainsonii; photo credit Nick Hanatake) Spurfowls. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
Figure
4.
Saturated (left) and bleached (right) throats in P. leucosceptus (above; photo credit Clayton Burneo) and P. swainsonii (below; photo credit Ricardo Santamaria).
3.2
Interspecific throat size differences and sexual size dimorphism
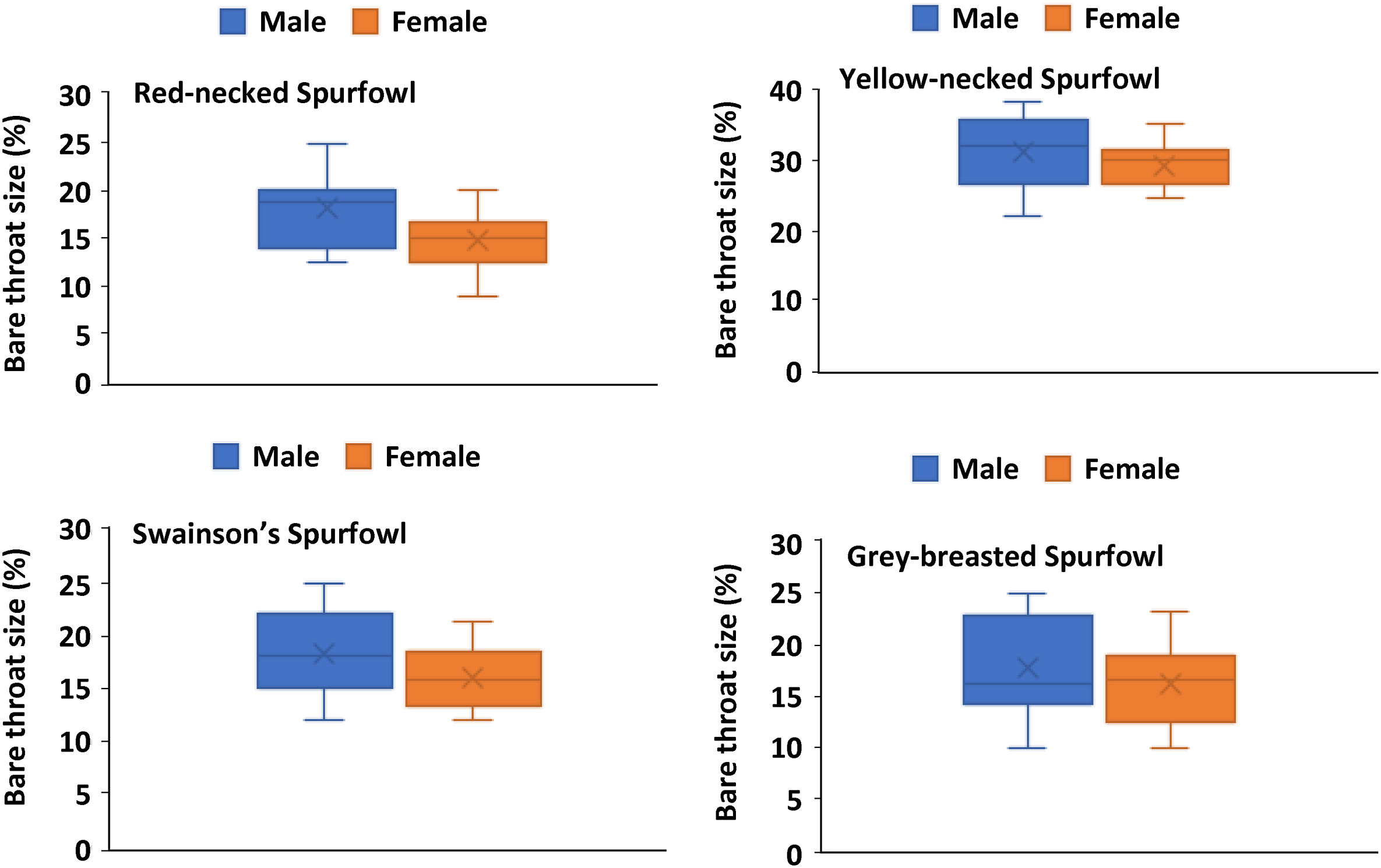
Among male spurfowls, the Yellow-necked Spurfowls have the largest throat area relative to the neck area. The same pattern was evident among their females. In other words, in Yellow-necked Spurfowl, less of the neck is covered by feathers (Table 5; Appendix Table S5). The percentage area of the bare throats of males was larger than that of the females in all four species (Fig. 5). However, statistically, the difference was only significant in Red-necked Spurfowl (H = 5.875, p = 0.0153, n = 50). For Swainson's Spurfowl, it was not significant (H = 0.1154, p = 0.734, n = 65), neither for Yellow-necked Spurfowl (H = 0.915, p = 0.338, n = 46) or Grey-breasted Spurfowl (H = 0.036, p = 0.848, n = 34) (Appendix Table S5) (see Fig. 6).
Table
5.
Using Two-way ANOVA, comparing throat sizes among the males and females of all four bare-throated spurfowls.
Figure
6.
Courtship display of Grey-breasted Spurfowl (P. rufopictus; photo credit John Bjorman) on the left and Cape Spurfowl (P. capensis; photo credit Robert Bochneck), a feather-throated spurfowl, on the right.
The spurfowl male performs frontal displays by rising above the female in front of him. Sometimes she runs away from him, and he follows behind her in this posture, but eventually, he manoeuvres himself in front, and then she looks at his breast feathers or upwards toward the head. During the high-rise posture, they extend their wings out from the body. The Bare-throated Spurfowl males rise significantly higher above their females than the Feather-throated Spurfowl males (H = 10.29, p = 0.001, n = 21). However, unlike the other Bare-throated Spurfowl males, the Swainson's Spurfowl male does not stretch his bare throat in a high arch above the females during courtship (van Niekerk, 1983). Notably, the Bare-throated Spurfowl males do not show a broader body in front of their females than the Feather-throated Spurfowl males (H = 1.017, p = 0.313, n = 19; Table 2) during courtship. The difference only lies in the height of the display.
3.4
Tree canopy
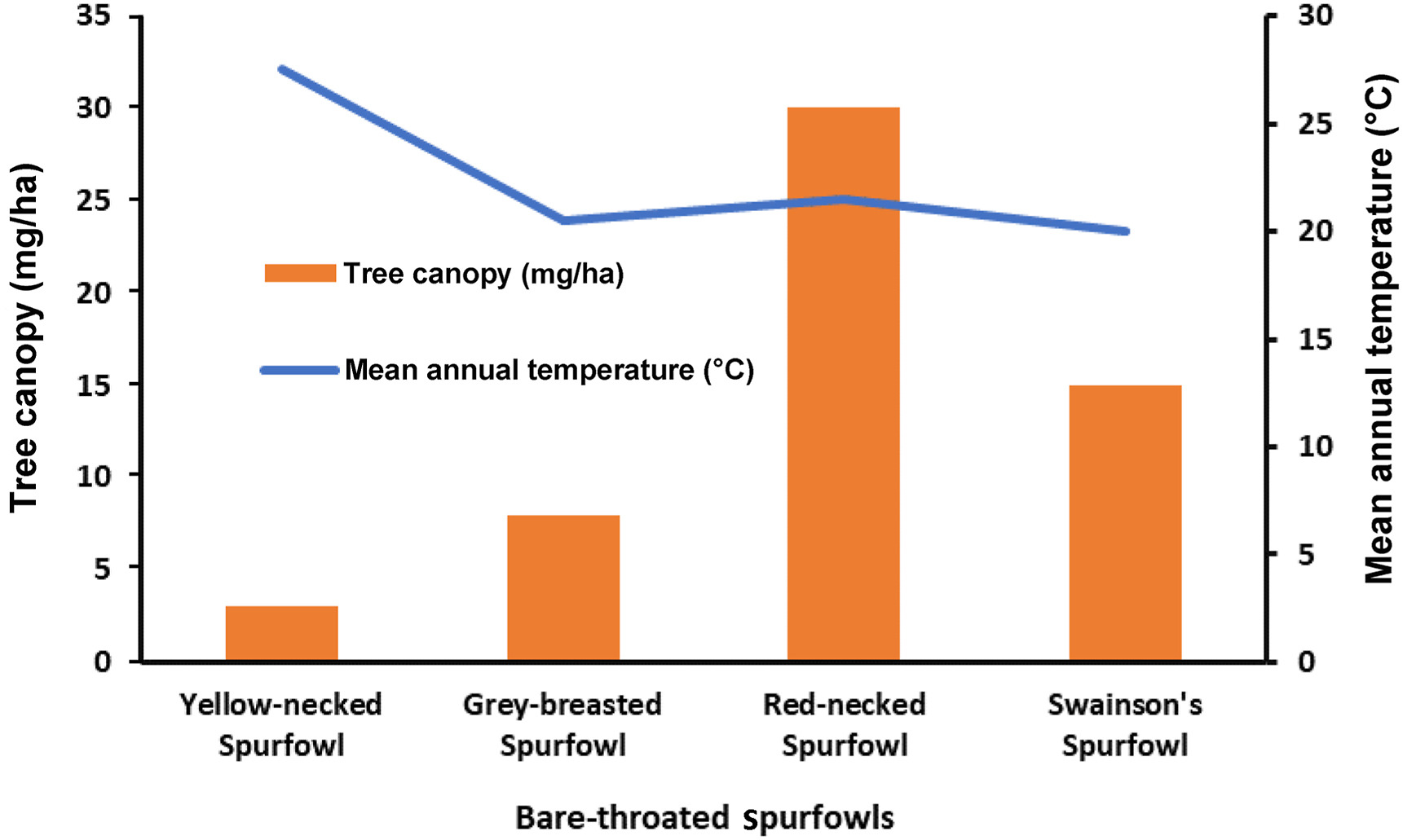
Yellow-necked Spurfowl lives in the sparsest area in terms of tree canopy (2–4 mg/ha) and the hottest environment, followed by Grey-breasted Spurfowl (6–10 mg/ha), Swainson's Spurfowl (10–20 mg/ha) and then Red-necked Spurfowl (20–40 mg/ha) (Fig. 7).
Figure
7.
Ambient mean temperatures and tree canopy density across the habitats in which the bare-throated spurfowls live.
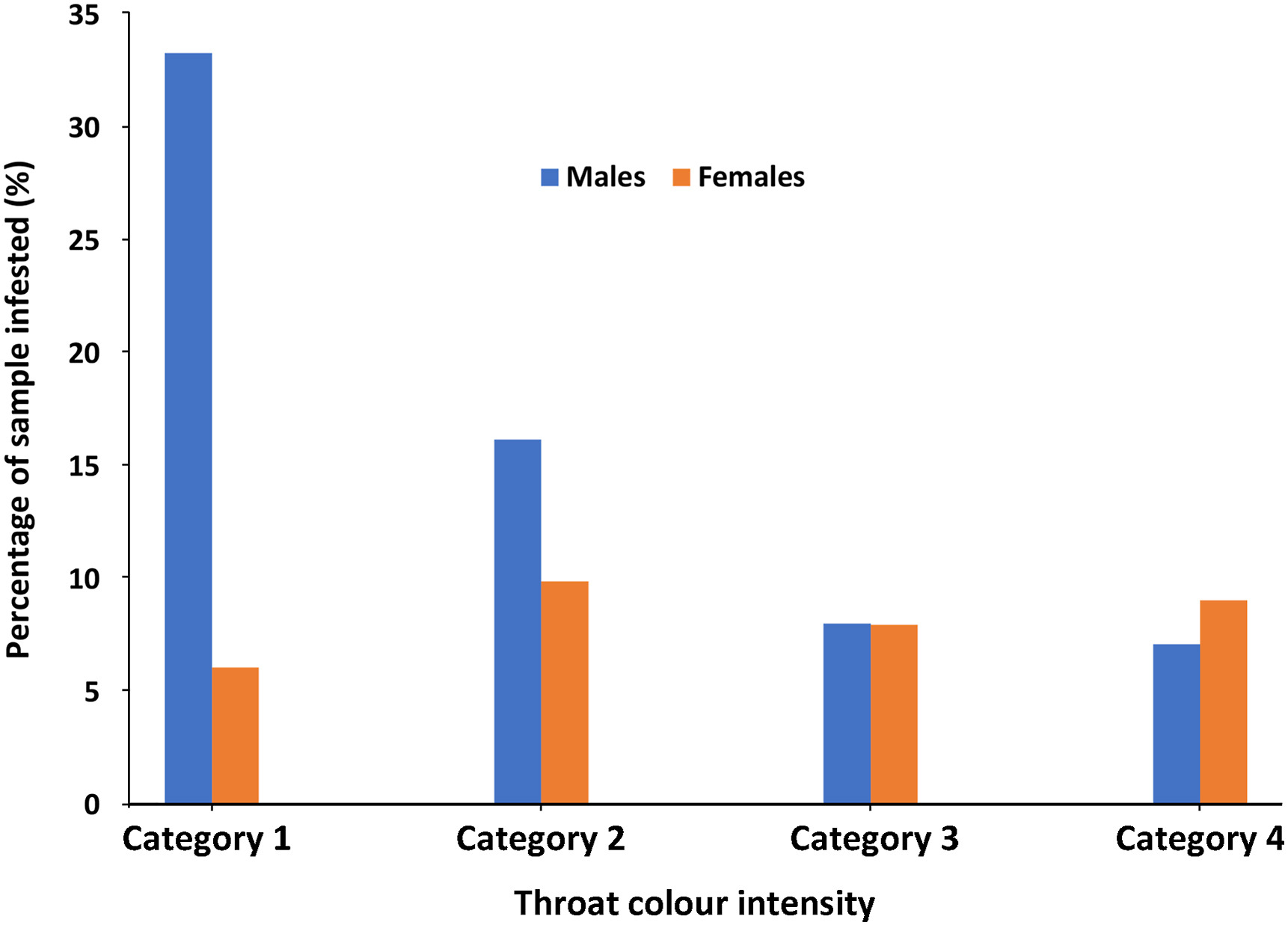
The bare throats of spurfowls were infested with ectoparasites, with 25% of all Yellow-necked Spurfowl infested, followed by Grey-breasted Spurfowl (9%), Red-necked Spurfowl (5%) and then Swainson's Spurfowl (1.7%) (Table 3: One-sample T-test; T = −15.199, p = 0.000618, n = 4). Most ectoparasites were ticks (Ixodidae) that often invaginated into the skin, causing a lesion. Overall, male birds with poor throat colours harboured most ectoparasites, whereas female throat infestation was more evenly spread across all four degrees of throat colour saturations (Fig. 8).
Figure
8.
Ectoparasite infestation of bare-throated spurfowl throats across different throat colour intensities. On a scale of 1–4, category 1 represents a bleached (low quality) throat colour and 4 represents a saturated throat colour (see Table 1). (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
Most spurfowl species do not have bare throats probably because they live in denser bush vegetation, riparian vegetation or forest edges (Madge and McGowan, 2001). The lack of tree canopy (shade) seems to relate to the development of bare throats in African spurfowl. Bare-throated spurfowls, on the other hand, live in more arid open vegetation. Most noticeable among these are the Yellow-necked Spurfowls with the largest bare throats. Their large throats are probably adaptations to hot climates with limited tree canopy cover in northeast Africa to maximise the dissipation of high brain temperature through the arteries in the throat skin (cf. van Niekerk et al., 2022). Grey-breasted Spurfowls also have bare throats and live in similarly arid conditions adjacent to Yellow-necked Spurfowl. Red-necked Spurfowls live in wetter vegetated habitats with more tree canopy but venture out in the open away from the continuous tree canopy. Swainson's Spurfowls live in the bushveld with more tree/bush canopy (Little, 2016). The latter three bare-throated species seemingly require less bare throat for thermoregulation than Yellow-necked Spurfowl.
The different colours among the bare-throated species possibly emerged as prezygotic mechanisms to minimise cross-breeding (Rubtsov, 2015). This is congruent with the cladogram of Mandiwana-Neudani et al. (2019) (Appendix Fig. S1), suggesting that the distinct throat colour of arid-adapted Yellow-necked Spurfowl and Grey-breasted Spurfowl may have evolved independently to reinforce assortative mating between them and the Red-necked Spurfowl and Swainson's Spurfowl.
The prezygotic function of different throat colours explains why females have coloured throats. In other words, while females (except perhaps Swainson's Spurfowl females) do not use coloured throats for signalling, it has not lost its prezygotic function among all four species. The upshot is that the bare throat had a primary thermoregulatory role which evolved into an additional secondary reproductive function.
Prasetya et al. (2020) studied the saturation of male plumage colours across a geographical gradient in Australia. They found that colour saturation did not respond to rainfall or primary productivity but rather to temperature. This finding contrasts with my study in which spurfowl's throat colour saturation correlates with rainfall and breeding. These different findings could be a matter of other mechanisms underlying feathers and bare skin and demonstrate our lack of understanding (Olson and Owens, 2005).
Among three species, namely Yellow-necked, Red-necked and Grey-breasted Spurfowls, males use this feature to court females at the start of the breeding season when their throat colours saturate. In contrast, female throat colours do not saturate seasonally among these three species (cf. Delhey et al., 2010). Swainson's Spurfowl males and females brighten their throats during breeding. Here the saturated bare throat of the male is used mainly to advertise his territory and not to impress females, as it is evident that he does not stretch his throat in an arch above females during courting like the other bare-throated spurfowls (van Niekerk, 1983).
Males and females of Swainson's Spurfowl probably consume carotenoid foodstuffs during the breeding season. Still, females of the other three species focus mainly on invertebrates to attain breeding conditions (Hall et al., 2012), while their males consume carotenoids. While handling three Swainson's Spurfowl males in the field, the red colour from their throats rubbed off, suggesting that it is a carotenoid-based substance secreted through the skin for cosmetic purposes (cf. Chiale et al., 2012). Studies are required to determine the exact role of carotenoids in this species.
Although some females develop saturated coloured throats, it is unregulated (except the female of Swainson's Spurfowl), i.e., not seasonally driven (see Fig. 2). For these females, throats are not signals of fecundity. On the other hand, the colour-saturated throats of males have become a seasonally driven ornament for sexual attraction. In this case, the response of male throat colours to climatic conditions is a physiological mechanism implanted by sexual selection (Freeman-Gallant et al., 2014). Males use these colours to attract females to their territories, but it may also signal that he holds a territory rich in food supplies for their offspring (Heinsohn, 2005). Females' less saturated throat colours may be a mechanism to prevent other males from interloping and fertilising females in established territories. It may also be that saturated throats would be too costly during the female's reproductive cycle, such as attracting predators (Hernández et al., 2012).
Sexual selection is at work to differentiate the males of Yellow-necked, Red-necked and Grey-breasted Spurfowls from their females in terms of seasonal throat colour saturation mechanisms, but not in Swainson's Spurfowl. The saturation of the female Swainson's Spurfowl throats probably indicates a sound immune system attractive to males, which is a sign of fecundity. Indeed, the acquisition of colourful ornaments by females to attract males is not uncommon among birds. According to Freeman-Gallant et al. (2014), males select females with ornaments that signal fecundity.
The graphs in Fig. 2 suggest that three bare-throated species, except Red-necked Spurfowl, produce eggs just after good rainy months. The period just after rain is likely the optimal period to find adequate nesting cover and an abundance of food for offspring (cf. van Niekerk, 2017). In these three species, the saturation of the throat in males builds up just before egg laying and then fades as egg laying and parental care prevail. One or two months earlier male throat saturation is only necessary when males establish territories to attract and court females. Less rainfall after the breeding season with less carotenoid-based food reverses saturation. A continuation of a saturated throat would be redundant because the breeding season is over, and females must raise their offspring. In other words, males do not have to compete for the brightest throat throughout the year. However, it is not uncommon to see males with bright throats harassing females with offspring (trying to split the mother from her chicks) during the non-rainy season among the bare-throated spurfowls (JHvN, unpublished). Indeed, this study shows that saturation occurs throughout the year, albeit at a lower frequency during the dry season.
Ectoparasites seem to preferentially frequent bare throats on spurfowl. Even the semi-bare orange throats of Clapperton's Spurfowl (P. clappertoni) in a section of their range in northeast Uganda attract ectoparasites (JHvN, unpublished). The higher prevalence of ectoparasites on Yellow-necked Spurfowl may be related to more extensive home ranges. They possibly forage widely for food, exposing them to a higher volume of ectoparasites. Alternatively, since they have large bare throats for heat dissipation, a rich network of arteries is situated close to the skin surface, making it an attractive area to suck blood. Ectoparasites do not invade the bare orbital skins, which probably means these areas do not have a rich network of arteries below a thin skin. Spurfowl with bleached throats attracts more ectoparasites, possibly because these birds live in low-quality habitats, which do not promote colourful throats. In other words, ectoparasites may avoid skin with carotenoids. Reproductive males with saturated throat colours avoid infestation to attract females. According to Borgia and Collis (1989), females do not readily pair with birds that show parasitism, avoiding an adverse effect on their immune system.
Acquiring food-based carotenoids for signalling may be costly because developing the brightest colours for signalling suggests that the birds must invest metabolic and energetic resources to achieve it. Specifically, advertisement crowing, attracting females, and avoiding predators are related to a colourful throat and makes up for this cost. Ostensibly, environmental factors assist in offsetting the cost of possessing a bare throat. Unlike the fully feathered throated spurfowl, the bare-throated birds live in more open habitats. This canopy free habitat provides a better chance for predator surveillance; therefore, the bare-throated spurfowl can survive with a bright bare throat, unlike the fully feathered spurfowl in denser habitats. Rainfall is a direct extrinsic factor that seems essential for successful breeding. Rainfall triggers the production of carotenoid food. Specifically, rainfall promotes throat saturation, territorial displays, and egg laying in quick succession.
The bare-throated spurfowls are a closely related subgroup among the spurfowl taxonomically (cf. Hall, 1963; Mandiwana-Neudani et al., 2019), but as the present research indicates, this reflects in their behaviour. Courtship behaviour varies among all spurfowl species because of the impetus of the saturated throat colours of males. Bare-throated spurfowl males rise higher than feather-throated spurfowl males during courtship, demonstrating the importance of the bare throat as an ornament. During fieldwork, it was evident how females stared at males' throats during these displays (JHvN, unpublished). She stands directly in front of the male, looking upwards, suggesting that the colourful throat signifies a potentially fit partner for breeding. The bare throat also plays a role in sexual dimorphism. In at least one species, the Red-necked Spurfowl, the male's bare throat is significantly larger than the female's. It results in a larger saturated red throat that has become a billboard that attracts females, which may very well aid the promiscuous nature of this species (van Niekerk et al., 2009).
The taxonomy of African spurfowls is currently contentious (cf. Crowe et al., 2022). Taxonomy is important, but spurfowl behaviour and ecology deserve a tremendous push since we know little about these aspects. Such an approach will undoubtedly help raise taxonomists' understanding of spurfowl relationships to another level (Xiao et al., 2017).
Ethics statement
All the data gathered for this manuscript was non-invasive. The data were retrieved from photographs from the online Macaulay Library at Cornell University. Macaulay Library granted special permission to reproduce the photographs depicted in this manuscript.
Declaration of competing interest
The author declares that he has no competing interests.
Acknowledgements
The Macaulay Library at the Cornell Lab of Ornithology granted permission to duplicate the photographs used in this paper. I also thank the two anonymous reviewers. Finally, thanks go to Dr Giovanni Forcina and Klarissa van Niekerek for their assistance.
Alerstam, T., Hedenström, A., Åkesson, S., 2003. Long-distance migration: evolution and determinants. Oikos 103, 247-260.
Anderson, C.M., Gilchrist, H.G., Ronconi, R.A., Shlepr, K.R., Clark, D.E., Fifield, D.A., et al., 2020. Both short and long distance migrants use energy-minimizing migration strategies in North American herring gulls. Mov. Ecol. 15, 26.
Arlt, D., Pärt, T., 2008. Post-breeding information gathering and breeding territory shifts in northern wheatears. J. Anim. Ecol. 77, 211-219.
Baert, J., Stienen, E.W., Heylen, B., Kavelaars, M., Buijs, R.J., Shamoun-Baranes, J., et al., 2018. High-resolution GPS tracking reveals sex differences in migratory behaviour and stopover habitat use in the lesser Black-backed Gull Larus fuscus. Sci. Rep. 8, 5391.
Bauer, S., Gienapp, P., Madsen, J., 2008. The relevance of environmental conditions for departure decision changes en route in migrating geese. Ecology 89, 1953-1960.
Betts, M.G., Hadley, A.S., Rodenhouse, N., Nocera, J.J., 2008. Social information trumps vegetation structure in breeding-site selection by a migrant songbird. Proc. R. Soc. A B 275, 2257-2263.
Bogdanova, M.I., Daunt, F., Newell, M., Phillips, R.A., Harris, M.P., Wanless, S., 2011. Seasonal interactions in the black-legged kittiwake, Rissa tridactyla: links between breeding performance and winter distribution. Proc. Biol. Sci. 278, 2412-2418.
Burger, J., Gochfeld, M., Kirwan, G.M., Garcia, E.F.J., 2020. Black-tailed gull (Larus
crassirostris), version 1.0. In: Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., Juana, E. (Eds.), Birds of the World. Cornell Lab of Ornithology, Ithaca, NY.
Ciaglo, M., Calhoun, R., Yanco, S.W., Wunder, M.B., Stricker, C.A., Linkhart, B.D., 2021. Evidence of postbreeding prospecting in a long-distance migrant. Ecol. Evol. 11, 599-611.
Clark, D.E., Koenen, K.K.G., Whitney, J.J., MacKenzie, K.G., DeStefano, S., 2016. Fidelity and persistence of ring-billed (Larus delawarensis) and herring (Larus argentatus) gulls to wintering sites. Waterbirds 39, 220-234.
Drent, R.H., 2006. The timing of bird's breeding season: the Perrins hypothesis revisited, especially for migrants. Ardea 94, 305-322.
Dwyer, T., 1972. An adjustable radio-package for ducks. Bird-Banding 43, 282-284.
Egevang, C., Stenhouse, I.J., Phillips, R.A., Petersen, A., Fox, J.W., Silk, J.R.D., 2010. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proc. Natl. Acad. Sci. USA 107, 2078-2081.
Fayet, A.L., Freeman, R., Anker-Nilssen, T., Diamond, A., Erikstad, K.E., Fifield, D., et al., 2017. Ocean-wide drivers of migration strategies and their influence on population breeding performance in a declining seabird. Curr. Biol. 27, 3871-3878.
Frederiksen, M., Descamps, S., Erikstad, K.E., Gaston, A.J., Gilchrist, H.G., Grémillet, D., et al., 2016. Migration and wintering of a declining seabird, the thick-billed murre Uria lomvia, on an ocean basin scale: conservation implications. Biol. Conserv. 200, 26-35.
Friesen, V., Burg, T., McCoy, K., 2007. Mechanisms of population differentiation in seabirds. Mol. Ecol. 16, 1765-1785.
Fromant, A., Bost, C.A., Bustamante, P., Carravieri, A., Cherel, Y., Delord, K., et al., 2020. Temporal and spatial differences in the postbreeding behaviour of a ubiquitous Southern Hemisphere seabird, the common diving petrel. R. Soc. Open Sci. 7, 200670.
Gilg, O., Strøm, H., Aebischer, A., Gavrilo, M.V., Volkov, A.E., Miljeteig, C., et al., 2010. Post-breeding movements of northeast Atlantic ivory gull Pagophila eburnea populations. J. Avian Biol. 41, 532-542.
González-Solís, J., Croxall, J.P., Oro, D., Ruiz, X., 2007. Trans-equatorial migration and mixing in the wintering areas of a pelagic seabird. Front. Ecol. Environ. 5, 297-301.
Hake, M., KjelleÂn, N., Alerstam, T., 2001. Satellite tracking of Swedish ospreys Pandion haliaetus: autumn migration routes and orientation. J. Avian Biol. 32, 47-56.
Hallworth, M., Marra, P., 2015. Miniaturized GPS tags identify non-breeding territories of a small breeding migratory songbird. Sci. Rep. 5, 11069.
Higuchi, H.O., Zaki, K., Fujita, G., Minton, J., Ueta, M., Soma, M., et al., 1996. Satellite tracking of white-naped crane migration and the importance of the Korean demilitarized zone. Conserv. Biol. 10, 806-812.
Higuchi, H., Shibaev, Y., Minton, J., Ozaki, K., Surmach, S., Fujita, G., et al., 1998. Satellite tracking of the migration of the Red-crowned crane Grus japonensis. Ecol. Res. 13, 273-282.
Kanai, Y., Sato, F., Ueta, M., Minton, J., Higuchi, H., Soma, M., et al., 1997. The migration routes and important restsites of Whooper Swans satellite-tracked from northern Japan. STRIX 15, 1-13.
Kazama, K., Tomita, N., Ito, M., Niizuma, Y., Takagi, M., Watanuki, Y., 2008. Responses in breeding behaviour of the Black-tailed Gull (Larus crassirostris) to different marine environments. In: Okada, H., Mawatari, S.F., Suzuki, N., Gautam, P. (Eds.), The Origin and Evolution of Natural Diversity. Hokkaido University, Sapporo, pp. 215-220.
Kassara, C., Evangelidis, A., Tsiopelas, N., Barboutis, C., Giokas, S., 2022. Seasonal and daily activity patterns by Eleonora's Falcon Falco eleonorae based on GPS telemetry: a contribution to the species' movement ecology at its breeding grounds. Bird Conserv. Int. 32, 154-171.
Kazama, K., Hirata, K., Yamamoto, T., Hashimoto, H., Takahashi, A., Niizuma, Y., et al., 2013. Movements and activities of male Black-tailed Gulls in breeding and sabbatical years. J. Avian Biol. 44, 603-608.
Klaassen, R.H.G., Ens, B.J., Shamoun-Baranes, J., Exo, K.M., Bairlein, F., 2012. Migration strategy of a flight generalist, the lesser black-backed gull Larus fuscus. Behav. Ecol. 23, 58-68.
Langendoen, T., Mundkur, T., Nagy, S., 2021. Flyway Trend Analyses Based on Data From the Asian Waterbird Census From the Period of 1987-2020. Wetlands International, Wageningen.
Lee, W.S., Kwon, Y.S., Yoo, J.C., Song, M.Y., Chon, T.S., 2006. Multivariate analysis and self-organizing mapping applied to analysis of nest-site selection in Black-tailed Gulls. Ecol. Model. 193, 602-614.
Liu, Y., Wu, H.D., 2017. Sea ice in the Bohai Sea and the northern Yellow Sea. Mar. Forecasts 34, 94-101 (in Chinese).
Lu, X., Zheng, G.M., 2003. Reproductive ecology of Tibetan Eared Pheasant Crossoptilon harmani in scrub environment, with special reference to the effect of food. Ibis 145, 657-666.
Marchant, S., Higgins, P.J., 1990. Handbook of Australian, New Zealand and Antarctic Birds. Oxford University Press, Oxford.
Mills, K.J., Patterson, B.R., Murray, D.L., 2006. Effects of variable sampling frequencies on GPS transmitter efficiency and estimated wolf home range size and movement distance. Wildl. Soc. Bull. 34, 1463-1469.
Mellone, U., Lopez-Lopez, P., Liminana, R., Urios, V., 2013. Summer pre-breeding movements of Eleonora's Falcon Falco eleonorae revealed by satellite telemetry: implications for conservation. Bird Conserv. Int. 23, 487-494.
Mizota, C., 2009. Nitrogen isotopic patterns of vegetation as affected by breeding activity of Black-tailed Gull (Larus crassiostris): a coupled analysis of feces, inorganic soil nitrogen and flora. Appl. Geochem. 24, 2027-2033.
Newton, I., 2008. The Migration Ecology of Birds. Academic Press, London.
Newton, I., 2011. Migration within the annual cycle: species, sex and age differences. J. Ornithol. 152, 169-185.
Rayner, M.J., Hauber, M.E., Steeves, T.E., Lawrence, H.A., Thompson, D.R., Sagar, P.M., et al., 2011. Contemporary and historical separation of transequatorial migration between genetically distinct seabird populations. Nat. Commun. 2, 332.
R Core Team, 2019. R: a language and environment for statistical computing. R Found. Stat. Comput. .
Rosana, A.M., Teresa, M., Ginés, V., Jacob, G.S., 2013. Pre-breeding ecophysiology of a long-distance migratory seabird. J. Exp. Mar. Biol. Ecol. 443, 162-168.
Rubolini, D., Massi, A., Spina, F., 2002. Replacement of body feathers is associated with low pre-migratory energy stores in a long-distance migratory bird, the barn swallow (Hirundo rustica). J. Zool. 258, 441-447.
Ryan, P.G., Petersen, S.L., Peters, G., Grémillet, D., 2004. GPS tracking a marine predator: the effects of precision, resolution and sampling rate on foraging tracks of African penguins. Mar. Biol. 145, 215-223.
Salewski, V., Bruderer, B., 2007. The evolution of bird migration - a synthesis. Naturwissenschaften 94, 268-279.
Shaffer, S.A., Tremblay, Y., Weimerskirch, H., Scott, D., Thompson, D.R., Sagar, P.M., et al., 2006. Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc. Natl. Acad. Sci. USA 103, 12799-12802.
Shamoun-Baranes, J., van Loon, E.E., Purves, R.S., Speckmann, B., Weiskopf, D., Camphuysen, C.J., 2012. Analysis and visualization of animal movement. Biol. Lett. 8, 6-9.
Shamoun-Baranes, J., Burant, J.B., van Loon, E.E., Bouten, W., Camphuysen, C.J., 2017. Short distance migrants travel as far as long distance migrants in lesser black-backed gulls Larus fuscus. J. Avian Biol. 48, 49-57.
Studds, C.E., Kyser, T.K., Marra, P.P., 2008. Natal dispersal driven by environmental conditions interacting across the annual cycle of a migratory songbird. Proc. Natl. Acad. Sci. USA 105, 2929-2933.
Vähätalo, A., Rainio, K., Lehikoinen, A., Lehikoinen, E., 2004. Spring arrival of birds depends on the north atlantic oscillation. J. Avian Biol. 35, 210-216.
van Doren, B.M., Conway, G.J., Phillips, R.J., Evans, G.C., Roberts, G.C.M., Liedvogel, M., et al., 2021. Human activity shapes the wintering ecology of a migratory bird. Global Change Biol. 27, 2715-2727.
Vega, M.L., Willemoes, M., Thomson, R.L., Tolvanen, J., Rutila, J., Samaš, P., et al., 2016. First-time migration in juvenile Common Cuckoos documented by satellite tracking. PLoS One 11, 12.
Wang, L., Han, J.K., Huang, M.P., 1991. The breeding ecology of Black-tailed Gulls (Larus crassiostris). Chin. J. Wildl. 3, 29-30 (in Chinese).
Weimerskirch, H., Delord, K., Guitteaud, A., Phillips, R.A., Pinet, P., 2015. Extreme variation in migration strategies between and within wandering albatross populations during their sabbatical year, and their fitness consequences. Sci. Rep. 5, 8853.
Zhang, S.W., Fan, Q.D., Zhao, F., Sun, W.L., Li, W.Q., Zhong, H.B., 2000. Observation on the breeding ecology of black-tailed gull (Larus crassiostris). J. Shandong For. Sci. Tech. 4, 14-16 (in Chinese).
Table
1.
Weighted scores were assigned to colour pigment intensity of the bare throat of the bare-throated spurfowls (see Appendix Table S1, S2, S3, S4).
Table
2.
Width and height of males and females of both bare-throated and feather-throated spurfowls converted to percentage differences between the two sub-groups of spurfowl (see materials and methods).
Table
4.
Correlation statistics between monthly throat colour saturation vs rainfall and monthly throat colour saturation vs. nest records of four bare-throated spurfowl species.

 DownLoad:
DownLoad:








 Email Alerts
Email Alerts RSS Feeds
RSS Feeds