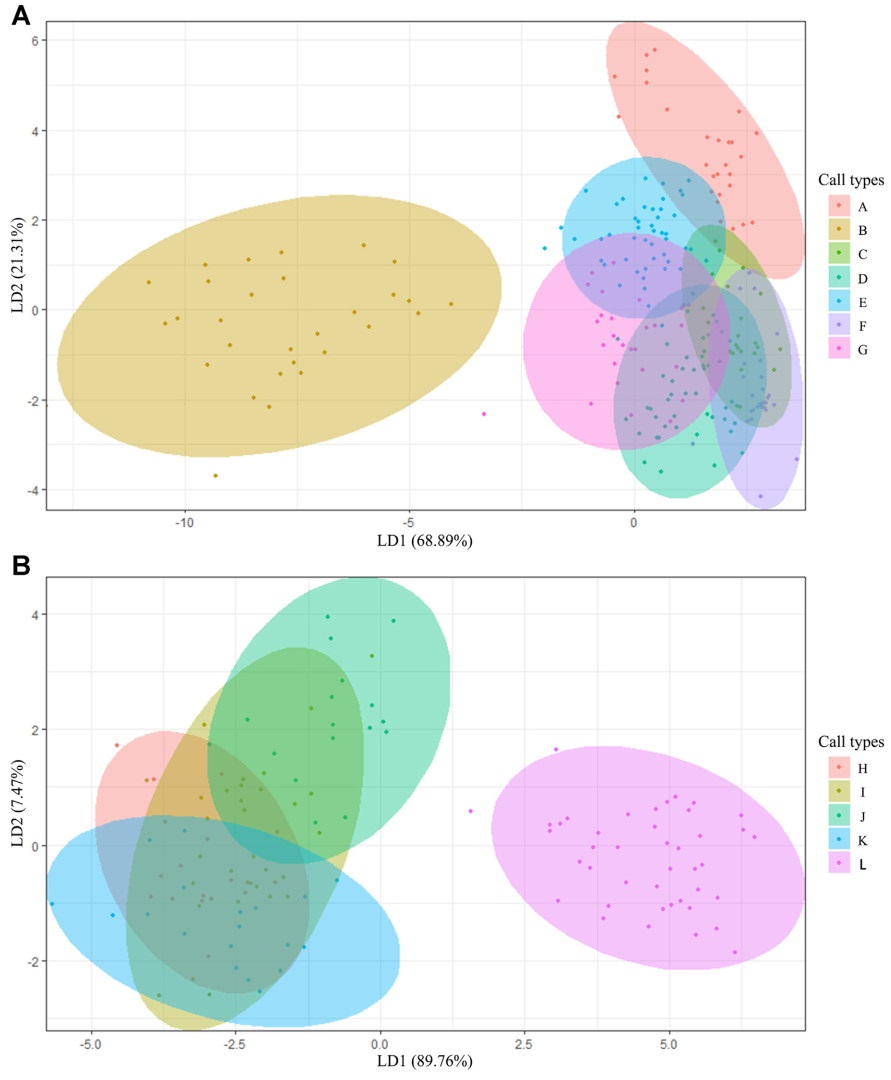
Signals within animals' vocal communication are considered functional referential and context-specific. Even in the absence of the context, receivers are expected to acquire the information of calls and respond specifically. Whereas the framework was supported by plenty of evidence, its exhaustivity in describing all animal vocalisations has been questioned. Here, we investigated the vocal repertoire of a cooperatively breeding species, Azure-winged Magpie (Cyanopica cyanus), to present evidence for referential signals. The results showed that Azure-winged Magpies had a relatively large vocal repertoire, consisting of twelve distinct calls. These calls were associated with the context including movement, begging for food, contact, vigilance against predators, etc. However, even the predator-specific alarm calls would induce various responses of receivers. This implies that multiple pieces of information are involved in the vocalisation, which could be utilised by the receiver to select an appropriate response based on the surroundings. Our study gives a detailed description of the context and function of the vocal repertoire in Azure-winged Magpies, laying the foundation for further investigation on the developmental mechanisms of bird vocalisations. This study also suggests that the referential signals of animal vocalisations may not be limited to the context-specific responses of receivers and need to be discussed from a broader perspective.
Identifying migration routines and the stopover sites of shorebirds is critical for understanding their life cycle and for conservation management (Battley et al., 2012; Giunchi et al., 2019). Such a task is essential for the East Asian–Australasian Flyway (EAAF), which is the most species-rich flyway globally but also has the most species in decline due to the loss of natural wetlands over the past several decades (Kirby et al., 2008; Murray et al., 2014; Studds et al., 2017).
Tracking the annual cycle of shorebirds can uncover their migration strategy, reveal potential stopover sites, and even show some behaviour adaptive to the natural tidal flat lost in the flyways (Lisovski et al., 2016; Piersma et al., 2021). Many efforts have been made on the EAAF, focusing mainly on long-distance migration or threatened species. For example, tracking the Great Knots (Calidris tenuirostris) provided further evidence that they used the Yellow Sea as the major stopover sites on both northward and southward migration and showed some short-stay sites probably for rest only before arriving at the Yellow Sea (Ma et al., 2013; Lisovski et al., 2016). The tracking of the Red Knot (Calidris canutus piersmai) showed that 'long jumpers' behaved as 'skippers'. Rather than staging at a single site for multiple weeks halfway during migration, they made several stops of up to a week. Recent reductions in the extent of suitable habitat in the Yellow Sea area (Murray et al., 2014; Piersma et al., 2016) may have contributed to the current pattern (Piersma et al., 2021). Tracking of Whimbrel (Numenius phaeopus) indicated that different populations have different reliance on the Yellow Sea, which might be related to their different population trends (Kuang et al., 2020).
However, less effort has been put into tracking the annual cycle of short-distance or least concerned species, despite their greater importance in ecosystem function and service provision (Inger et al., 2015). The neglect of common species hinders our knowledge of their migration route, breeding/wintering area, and the potential risk they may face during their life cycle. Population collapse can occur even in species with a billion individuals (Bucher, 1992).
Pied Avocets (Recurvirostra avosetta) are common shorebirds that occur across most regions of Asia, Europe, and Africa (IUCN, 2020). Males and females do not show strong sexual size dimorphism, and the breeding system is seasonally monogamous, with both sexes performing egg incubation and chick brooding (Pierce et al., 2020). Historically, Pied Avocets are common breeding waterbirds on the east coast of England during the early 19th century but disappeared in the mid-19th century due to the loss of natural habitat and overharvest of their eggs by people. They were reintroduced one hundred years later with the reconstruction of habitats and other methods in the 1940s (Hill, 1988). In the EAAF, the population of Pied Avocets is estimated to be 100, 000 (Wetlands International, 2020). Although the overall population trend is not clear, a local study found that the winter peak in Shenzhen Bay/Deep Bay in southern China has been decreasing in recent years (Choi et al., 2020), which means we need a focus on this common but neglected species.
The migration of the Pied Avocet has not been well studied, although some migration routes were revealed by banding in Europe (Hotker and West, 2005; Chambon et al., 2018), which is especially poorly documented in Asia. Their traditional breeding range was distributed from Central Asia to Southeast Siberia and Northeast China. However, in recent years, Pied Avocets have been documented breeding in areas farther south, for example, in the Bohai Bay of the Yellow Sea (Lei et al., 2021), in Chongming Dongtan in the East China Sea (Wanjuan Ke, personal communication), and in Hong Kong (Allcock et al., 2012).
Tracking avocets can also provide insight into sex-specific migration strategies. The migration timing and wintering area are different between sexes and are common in shorebirds. Migration timing (arrival at or departure from breeding/wintering grounds) is supposed to relate to the breeding system. Males usually arrive at the breeding area earlier than females in shorebirds to occupy the territory. Males that do not provide incubation or brooding leave earlier from the breeding area than females (Meissner, 2006). Males and females have different winter distributions that are supposed to be related to sexual size dimorphism (Nebel, 2005) or resource partitioning (Duijns et al., 2014). Studying the sex differences in shorebirds in terms of migration strategies can be helpful for detecting the factors linked to intraspecific variation, explaining their distribution pattern, and finding conservation gaps.
The wetlands of eastern China support a large non-breeding population of Pied Avocets (Cao et al., 2009). However, to the best of our knowledge, the annual routines and critical stopover sites of Pied Avocets have not been revealed. In this study, we tracked the avocets breeding in the Yellow Sea using satellite transmitters. We aim to identify their annual routines and stopover sites. We also explored sex-differential migration in this species. We will advance our understanding of migration ecology and fill the conservation knowledge gap for a common shorebird species in the EAAF.
2.
Methods
2.1
Study area and data collection
We captured Pied Avocets (hereafter "avocets") at two sites in northern Bohai Bay, Yellow Sea: (ⅰ) Nanpu saltpans, and (ⅱ) Yuetuo Island, both in Tangshan, Hebei Province, China (Fig. 1). Nanpu saltpans (118.1–118.3° E, 39.0–39.2° N) are an artificial habitat for waterbirds in the EAAF (Lei et al., 2018). Yuetuo Island (118.8–118.9° E, 39.1–39.2° N) is an artificial sand island located 50 km east of the Nanpu saltpans. More than 500 breeding pairs were recorded at the study sites (Wu, 2019–2021, unpublished results). Avocets arrived at the study sites in early March and started breeding in late April (Lei et al., 2021). They were captured using walk-in traps (0.8 m × 0.8 m × 0.6 m) or clap nets (N2019 = 27, N2020 = 13) during the incubation period.
Figure
1.
Annual distribution of tracked Pied Avocets. The heatmap represents low densities to high densities of locations from pale to dark.
Captured individuals were banded with a metal ring on the left tibia and colour flags (blue over yellow) on the right tibia with a unique code. A blood sample of 10–80 μL was collected from the brachial vein with a cotton swab and stored in a centrifuge tube at −20 ℃ for subsequent molecular sexing (Van der Velde et al., 2017).
In 2019–2020, we captured 40 adult avocets (22 males, 18 females) and outfitted them with tracking devices using a leg-loop harness (Verhoeven et al., 2020). Two types of transmitters were used: (1) HQPG2009P (N = 38, 9 g solar GPS/GSM, Hunan Global Messenger Technology Ltd., Changsha City, China) was programmed to record one location every 2–12 h, and (2) HQPG 1206 (N = 2, 7 g solar GPS/GSM) was programmed to record one location every 3–12 h.
To understand the present situation of critical stopover sites (see Results), we visited Lianyungang city, Jiangsu Province in 2020 to search for the 17 tagged avocets known to be present there during that autumn. In addition, we counted avocets from 31 October to 1 November in Lianyungang.
2.2
Tracking data
We retrieved the tracking data via the service platform of HQXS (http://www.hqxs.net), and the locations of transmitters with accuracy classes A, B, and C were retained (representing precisions of 5 m, 10 m, and 200 m, respectively).
Location was defined as the GPS point of transmitters; site was defined as the area of a bird staged for a while. Two avocets that lost signal within three months after release were excluded. Two avocets lost their signals in November 2019 during their southward migration. From July 2019 to June 2021, 36 individuals completed at least a southward or northward migration episode (single trip from wintering to breeding area or from breeding to wintering area, Appendix Table S1). There were 46 complete northward episodes from 35 individuals (19 males, 16 females) and 47 complete southward episodes from 35 individuals (18 males, 17 females).
Many avocets had second brood in southern Bohai Bay and Laizhou Bay of Bohai sea, after their reproduction in northern Bohai Bay. Thus, we take the farthest area south of the Bohai Sea (36.6° N) as the southern boundary of the breeding area. Most individuals would not travel long distances after arriving at the middle and lower reaches of the Yangtze River. Thus, we take the northern boundary of the middle and lower reaches of the Yangtze River (31° N) as the northern boundary of the wintering area. Based on the annual movements of the tracked avocets, we defined that the breeding and wintering areas were above 36.6° N and below 31° N, respectively. We considered an individual to be stationary when the recorded transmitter showed a speed of ~0 km/h (Zhu et al., 2021). Thus, the remaining stationary locations fell between these two latitudes and were considered stopping areas (Chan et al., 2019). The heatmap was used to describe the avocet breeding, wintering, and stopping areas in QGIS 3.20.0. Based on geographical location, we categorized the habitat of stationary locations into two types: (ⅰ) inland wetlands, and (ⅱ) coastal wetlands.
During migration, we assumed that the last sites where birds remained within 50 km for more than 48 h were considered stopover sites (Kuang et al., 2020). The departure and arrival dates at the breeding area and wintering areas were determined by their last site. During southward migration, the last site in the breeding area with a duration of staying more than 48 h within 50 km was treated as the start of southward migration. The time of the last location on this site was the departure date. If tracked individuals left their breeding area and then flew back, the last time was used as the departure time. The first site in the wintering area with a duration of staying more than 48 h within 50 km was treated as the last site of southward migration. The time of the first location on this site was the arrival date. When the migration tracking signal was lost over 24 h but less than 10 d, the starting date of migration was defined as the medium date between the date of the last location in the breeding area and the first location during migration. Individuals whose signal was lost over 10 d were excluded. The departure and arrival dates of northward migration were calculated similarly.
We used the R package 'geosphere' (Hijmans et al., 2017) to calculate the distance between the last location of all breeding/stopover sites before migration and the first location when arriving at the next stopping/wintering sites. The migration distance is the sum of the distance between the breeding sites, wintering sites, and all stopover sites on the route.
Tracked individuals with more than one repeated southward/northward migration episode (and individuals with incomplete episodes but had locations in breeding areas or wintering areas for more than one year) were collected to calculate breeding/wintering site fidelity at the individual level. We distinguished birds wintered in inland wetlands or coastal wetlands. Individuals were considered to have changed their wintering area only when they changed their wintering sites from inland wetlands to coastal wetlands or vice versa. Wintering site fidelity was determined by the wintering area change percentage between two years. A lower percentage indicates higher site fidelity. The breeding site fidelity was calculated similarly.
2.3
Statistical analysis
The linear mixed model (LMM) from the R package 'lme4' (Bates et al., 2015) was used to explore whether migration distance or duration differed between sexes and seasons, with individuals as random effects. We also used LMM to explore whether departure and arrival times of southward and northward migration differed between the sexes and years, with individuals as random effects. The statistical significance of fixed effects was calculated with the R package 'lmerTest' (Kuznetsova et al., 2017). Based on the wintering location of avocets, Fisher's exact test was used to explore whether males and females differed in the winter distribution. The t-test was used to explore whether the arrival and departure times of Lianyungang during northward migration differed between sexes. Analysis was conducted in R (R Core Team, 2020). Data are presented as the mean ± standard deviation (SD).
3.
Results
3.1
Annual routines, wintering and stopover sites
During southward migration, most individuals departed from Bohai Bay, but one departed from Liaodong Bay, Liaoning Province, in 2019 and 2020, and one departed from Hulun Buir, the Inner Mongolia Autonomous Region, in 2020 (Figs. 1 and 2), because they moved northwards after breeding in Bohai Bay. Tracked birds wintered in two areas of southern China: inland wetlands in the middle and lower reaches of the Yangtze River (hereafter "MLYR") and coastal wetlands in southeastern China (Figs. 1 and 2). Most individuals chose MLYR as their wintering sites (22 males, 7 females), and other individuals wintered in coastal wetlands (4 males, 8 females). Seven (all females) wintered in both areas and shifted from inland wetlands to coastal wetlands in the same season. There was a significant difference between sexes in the winter distribution. Males prefer inland wetlands, and females prefer coastal wetlands (p = 0.001; Appendix Table S1). In addition, one individual wintered on Taiwan Island in 2020, and two wintered in Yancheng coast wetland, Jiangsu Province, which is north of the wintering areas of other avocets. The coordinates of these areas are described in Fig. 1.
Figure
2.
Annual migration routes of Pied Avocets. The northernmost blue line represents individual H51. In 2020 and 2021, H51 bred in Hulun Buir, north of the Bohai Sea.
The change rates of both wintering area and breeding sites between years were small. In the 17 avocets with wintering tracking data both in 2019 and 2020 (Appendix Table S1), 14 wintered in inland areas, 3 wintered in a coastal area in 2019, and 11.76% of individuals (2 in 17) changed wintering areas in 2020 (both switched from inland to coastal wetlands). Twenty-one individuals captured in Bohai Bay in 2019 revisited their breeding sites in 2020. In 2021, 21 of 24 individuals (87.5%) were still bred at previous breeding sites. Thus, on average, 8.7% of individuals changed their breeding sites in three years.
During both southward and northward migration, most stopover sites were located along the coastal wetlands, while a few stopover sites were in the inland wetlands (Fig. 1).
The coastal wetland of Lianyungang (Lianyungang as follows) is the most important stopover site for avocets. More than 60% of tracked individuals staged in Lianyungang during southward migration, with an average of 36 days of stopover duration (Table 1). During the northward migration, more than 40% of tracked individuals staged in Lianyungang, with an average of 18 days of stopover duration. In the autumn of 2020, we surveyed Lianyungang and observed 39, 700 avocets on the tidal flat within two days, up to 40% of the population in the EAAF (Appendix Fig. S1).
Table
1.
The number of tracked Pied Avocets staged in Lianyungang during four migration periods.
Year
Migration period
Tracked individuals staged
Proportion of staged individuals (%)
Average duration of staging (d)
Range of duration (d)
2019
Southward
13
65
35.9 ± 34.5
1–110
2020
Southward
17
61
2020
Northward
9
41
18.4 ± 13.2
1–52
2021
Northward
12
52
Only individuals who staged for more than two days were calculated for the duration.
On average, avocets started southward migration on 23 October (Table 2). The majority reached the wintering grounds on 22 November. Males and females did not differ significantly in the departure time from the breeding sites (p = 0.59), the time of arrival at the wintering sites (p = 0.062), or the duration (p = 0.098). On average, avocets started northward migration on 22 March, and the majority reached the breeding sites on 7 April. The males departed from the wintering grounds earlier than females (p = 0.048), but both sexes did not differ significantly in the time of arrival at the breeding sites (p = 0.59). Thus, the duration of northward migration of males was slightly longer than the duration of northward migration of females but not significantly different (p = 0.126). For tracked birds staged in Lianyungang, males arrived at Lianyungang earlier than females (p = 0.013) in northward migration, but their departure time from here was similar (p = 0.88), which means that males stayed longer than females.
Table
2.
Estimated regression parameters, standard errors, profile confidence intervals (2.5% PCL and 97.5% PCL), χ2 values, df, and P values for each predictor variable to investigate the effects of season/year and sex (as fixed effects) on the migration distance, migration duration and migration timing of the Pied Avocet (Recurvirostra avosetta).
The great circle distances between the wintering and breeding sites ranged from 80 to 2350 km with an average of 1124 km, with no significant differences between males and females (p = 0.086; Table 2). However, the migration duration of males was longer than the migration duration of females (p = 0.048). Avocets migrated longer distances (p < 0.001) and shorter durations (p = 0.002) during northward migration (1256 ± 471 km and 15.61 ± 17.12 d) than southward migration (1019 ± 268 km and 30.13 ± 36.52 d).
4.
Discussion
4.1
Wide-range wintering area
Although the tracked birds were caught in a small breeding area (within 50 km) in Bohai Bay, they showed a broad wintering area with a maximum ca. 1000 km apart from each other, from the inland freshwater environments (MLYR) to coasts in southern China. Most individuals wintered in MLYR, while a few individuals overwintered in coastal wetlands. This result is inconsistent with those about only 15% (14, 590) and 32% (31, 971) of the estimated population of avocets overwintered on the inland wetlands at the beginning of this century (Cao et al., 2009) and in recent years (Tao et al., 2017), respectively. This difference may arise from the research method, i.e., the counting data of these two studies reflected the general wintering population, but the tracking data of this study were based on the breeding population in Bohai Bay, which may prefer the inland wintering area. Thus, more tracking data from other breeding areas or wintering areas are needed. In western Europe, most avocets tend to use coastal wetlands during the wintering period (Moreira, 1995; Hotker and West, 2005). Shorebirds that habituated both coastal and inland wetlands could also be found in Black-tailed Godwit (Limosa limosa; Estrella and Masero, 2010; Lourenço et al., 2010) and Dunlin (Calidris alpina; Abad-Gómez et al., 2020). Compared to coastal habitats, inland freshwater habitats where pathogens and parasites are generally more abundant may increase investment in the immune system (Piersma, 2003; Abad-Gómez et al., 2020). Avocets that spend more energy resisting pathogens may have a negative impact on subsequent migration and breeding. Female avocets may be more constrained by such energy costs than males; thus, they prefer wintering in coastal wetlands.
Winter site conditions substantially influence the survival of migratory species (McMahon et al., 2003; Woodworth et al., 2017). Dispersed wintering patterns may positively affect population survival by diluting the habitat degeneration risk of specific wintering areas (Cresswell, 2014; Bracey et al., 2018). However, avocet also showed winter site fidelity at the individual level and showed strong breeding site fidelity at the population level; thus, they are still sensitive to habitat loss in both wintering and breeding areas.
4.2
Migration strategy
Males and females did not differ in departure time from the breeding area, and arrival time at wintering grounds may be due to small sexual size dimorphism and both sexes participating in reproduction. Males depart from wintering areas earlier than females, but they did not differ in arrival time at breeding sites, which is in line with the result of Black-tailed Godwits (Zhu et al., 2021). This finding can be explained by males spending more time at stopover sites. Their arrival time at Lianyungang is earlier than the arrival time of females, but the departure time from Lianyungang is similar between the sexes.
The distance and main stopover sites of northward and southward migration routes are similar. The migration distance of avocets breeding in Bohai Bay is similar to some inexperienced individuals on the French Atlantic coast in a ringing study (Chambon et al., 2018). Compared to other common migration shorebirds in the EAAF (e.g., Bar-tailed Godwit Limosa lapponica, Red Knot, Great Knot, and Whimbrel) that fly over 10, 000 km (Lisovski et al., 2016; Kuang et al., 2020; Piersma et al., 2021), avocets can be classified as short-distance migration species. Furthermore, our results indicated that short-distance migratory species also rely on the stopover site during migration, which should not be neglected.
The migration distance of avocets ranged from 80 km to more than 2300 km, showing a 30-fold variation. Some individuals migrated from breeding areas to wintering areas or vice versa with a non-stop flight. Avocets can fly more than 1000 km within a single day, but only a small proportion of individuals use such a strategy. Overall, avocets showed great diversity in their migration behaviour. They can finish the migration without rest or spend over three months at stopover sites. The migration strategy of avocets can be described as a continuum, from the "cost minimization strategy" (for the individuals stopping for a long time in stopover sites) to the "time minimization strategy" (for the individuals with non-stop migration), and more research about such diversity strategies can fill the knowledge gap of individual specifications of shorebird migration.
4.3
Importance of Lianyungang
Both northward and southward migration routes showed the "umbrella" pattern, which highlighted the importance of Lianyungang in the migration path of the population of avocets in East Asia (Fig. 3). Our survey suggested that Lianyungang supported nearly 40% of the avocet population in the EAAF. Another report from Spoon-billed Sandpiper in China (2022) found more than 22, 000 avocets in Lianyungang during southward migration in the same year. This number may be underestimated because their count focuses on the Asian Dowitcher (Limnodromus semipalmatus). In addition to northward migration in 2020, more than half of the tracked avocets refuel in Lianyungang during southward migration (Table 1). Lianyungang is in the middle point between the breeding sites of Bohai Bay and the wintering sites of MLYRR, which have a straight-line distance of approximately 500 km. There was no large and suitable habitat for avocet after they migrated southward across the inland Shandong Peninsula until Lianyungang (Fig. 1). Lianyungang is the first staging site for avocets to migrate from Bohai Bay and is also the last refuelling site before they fly across the peninsula to breeding sites. If avocets choose a detour flight for an extra 500 km to fly along the coastline, their single non-stop migration distance increases from 500 km to 1000 km. In addition to avocets, Lianyungang supported over 1% of the flyway populations of other 21 shorebird species, and most are long-migration distance species threatened by losing important stopover sites (Chan et al., 2019). In particular, the Asian Dowitcher, for 97.5% of its estimated global population, was recorded in 2019 in Lianyungang (Yang et al., 2021). If Lianyungang is lost, there will be a serious impact on the annual migration of avocets, as well as other birds.
Figure
3.
Flocks of Pied Avocets in Lianyungang during northward migration. (Photo credits: Yongxiang Han).
Unfortunately, 27% of Lianyungang's intertidal soft-sediment habitats have been reclaimed from 2003 to 2018 (Chan et al., 2019), and a large area of tidal flat has been invaded and occupied by the salt-water cordgrass Spartina alterniflora. However, with only a provincial wetland park being established, Lianyungang still lacks adequate protection.
5.
Conclusions
This study is the first to examine the annual routines and main stopover sites of Pied Avocets in the EAAF. As a short-distance migratory species, avocets tracked in our study spent their whole annual life cycle within China, which is different from most shorebirds in the EAAF. They connected the inland freshwater wetlands and coastal wetlands in the Yellow Sea. Thus, their population fate is closely linked with the status of both types of wetlands in China. Identifying their critical stopover sites, such as Lianyungang, and building protected areas for the areas is vital for future conservation. The international responsibilities for conserving wetlands require China to balance economic development and biodiversity conservation. We strongly recommend that the coastal wetlands of Lianyungang be enrolled in protected areas and guaranteed under law as soon as possible.
Authors' contributions
YW and WL conceived the experiments, analyzed the data, wrote the manuscript, and reviewed the drafts; BZ analyzed the data and reviewed the drafts; JX performed the experiment in the field and analyzed the data; YM analyzed the data; ZZ conceived and designed the experiments, and reviewed drafts of the paper. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Zhengwang Zhang reports financial support was provided by National Natural Science Foundation of China and Non-profit Foundation of Marine Environment and Ecological Conservation of CNOOC. Weipan Lei reports financial support was provided by National Natural Science Foundation of China.
Acknowledgement
We thank volunteers (Chuyu Cheng, Shunlong Liu, Peichen Lin, Xiangwan Liu, Dan Li, Luoyi Kang, Qingxin Ma, Hengjun Xiao), colleagues from Beijing Normal University, and the Global Flyway Network who joined and offered their help in Tianjin and Nanpu. We also thanks Professor Weidong Kong, Dr. Hui Lin, Yang Sun and Jing Zhang for their advice to refine the paper. We also thank the two reviewers who provided valuable and insightful comments which greatly improved the manuscript. This research was funded by the National Natural Science Foundation of China (grant number 31830089, 32270518, 31801985), and the Non-profit Foundation of Marine Environment and Ecological Conservation of CNOOC (grant number CF-MEEC/TR/2020-20).
All experimental protocols of bird capture and handling were strictly complied with the requirement of Chinese Wild Animal Protection Law and all necessary permission for bird banding was obtained.
Bergman, T.J., Beehner, J.C., Cheney, D.L., Seyfarth, R.M., 2003. Hierarchical classification by rank and kinship in baboons. Science 302, 1234-1236.
Beynon, P., Rasa, O., 1989. Do dwarf mongooses have a language-warning vocalizations transmit complex information. S. Afr. J. Sci. 85, 447-450.
Blumstein, D.T., Armitage, K.B., 1997. Does sociality drive the evolution of communicative complexity? A comparative test with ground-dwelling sciurid alarm calls. Am. Nat. 150, 179-200.
Boucherie, P.H., Loretto, M.C., Massen, J.J., Bugnyar, T., 2019. What constitutes "social complexity" and "social intelligence" in birds? Lessons from ravens. Behav. Ecol. Sociobiol. 73, 1-14.
Brewer, D.E., 2022. The vocal repertoire of Blue Jays (Cyanocitta cristata): spectrographic snapshots and suggested nomenclature. Wilson J. Ornithol. 134, 309-317.
Catchpole, C.K., Slater, P.J., 2008. Bird Song: Biological Themes and Variations. Cambridge University Press, Cambridge.
Collier, K., Radford, A.N., Townsend, S.W., Manser, M.B., 2017. Wild dwarf mongooses produce general alert and predator-specific alarm calls. Behav. Ecol. 28, 1293-1301.
Conner, R.N., 1985. Vocalizations of common ravens in Virginia. Condor 87, 379-388.
Da, X.W., Xian, L.L., Luo, J.J., Gao, L.F., Du, B., 2018. Azure-winged magpies Cyanopica cyanus trade off reproductive success and parental care by establishing a size hierarchy among nestlings. Ibis 160, 769-778.
Delehanty, D.J., 2022. Raven control from a conservation biology perspective. Hum-Wildl. Interact. 15, 23.
Dutour, M., Walsh, S.L., Ridley, A.R., 2021. Australian magpies adjust their alarm calls according to predator distance. Bioacoustics 30, 458-468.
Engesser, S., Ridley, A.R., Townsend, S.W., 2017. Element repetition rates encode functionally distinct information in pied babbler 'clucks' and 'purrs'. Anim. Cogn. 20, 953-960.
Evans, C.S., 1997. Referential signals. In: Owings, D.H., Beacher, M.D., Thompson, N.S. (Eds.), Communication. Plenum Press, New York, pp. 99-143.
Fichtel, C., Kappeler, P.M., 2002. Anti-predator behavior of group-living Malagasy primates: mixed evidence for a referential alarm call system. Behav. Ecol. Sociobiol. 51, 262-275.
Fishbein, A.R., Idsardi, W.J., Ball, G.F., Dooling, R.J., 2020. Sound sequences in birdsong: how much do birds really care? Philos. T. Roy. Soc. B 375, 20190044.
Grieves, L.A., Logue, D.M., Quinn, J.S., 2015. Vocal repertoire of cooperatively breeding Smooth-billed Anis. J. Field Ornithol. 86, 130-143.
Harada, S., 1997. Age determination of the azure-winged magpie Cyanopica cyana (Aves) by moult patterns of alulae, some wing-coverts and rectrices. Raffles Bull. Zool. 45, 265-273.
Harcourt, A.H., Stewart, K.J., 1994. Gorillas' vocalizations during rest periods: signals of impending departure? Behaviour 130, 29-40.
Hope, S., 1980. Call form in relation to function in the Steller's Jay. Am. Nat. 116, 788-820.
Horn, L., Scheer, C., Bugnyar, T., Massen, J.J., 2016. Proactive prosociality in a cooperatively breeding corvid, the azure-winged magpie (Cyanopica cyana). Biol. Lett. 12, 20160649.
Komeda, S., Yamagishi, S., Fujioka, M., 1987. Cooperative breeding in azure-winged magpies, Cyanopica cyana, living in a region of heavy snowfall. Condor 89, 835-841.
Leighton, G.M., 2017. Cooperative breeding influences the number and type of vocalizations in avian lineages. Proc. R. Soc. B 284, 20171508.
Leighton, G.M., Birmingham, T., 2021. Multiple factors affect the evolution of repertoire size across birds. Behav. Ecol. 32, 380-385.
Macedonia, J.M., Evans, C.S., 1993. Essay on contemporary issues in ethology: variation among mammalian alarm call systems and the problem of meaning in animal signals. Ethology 93, 177-197.
Manser, M.B., 2001. The acoustic structure of suricates' alarm calls varies with predator type and the level of response urgency. Proc. R. Soc. B 268, 2315-2324.
Manser, M.B., 2016. Referents and semantics in animal vocalizations. In: Bee, M.A., Miller, C.T. (Eds.), Psychological Mechanisms in Animal Communication. Springer, Cham, pp. 223-249.
Marler, P., 1967. Animal communication signals: we are beginning to understand how the structure of animal signals relates to the function they serve. Science 157, 769-774.
Marler, P., 2008. Bird calls: a cornucopia for communication. In: Marler, P., Slabbekoorn, H. (Eds.), Nature's Music. Elsevier Academic Press, Cambridge, pp. 132-177.
Massen, J.J., Haley, S.M., Bugnyar, T., 2020. Azure-winged magpies' decisions to share food are contingent on the presence or absence of food for the recipient. Sci. Rep. 10, 16147.
Morton, E.S., 1977. On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. Am. Nat. 111, 855-869.
Oksanen, J., Blanchet, F., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al., 2019. vegan: Community Ecology Package. R package version 2.5-6. .
Owren, M.J., Rendall, D., 1997. An affect-conditioning model of nonhuman primate vocal signaling. In: Owings, D.H., Beacher, M.D., Thompson, N.S. (Eds.), Communication. Plenum Press, New York, pp. 299-346.
R Core Team, 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing. .
Raveling, D.G., 1969. Preflight and flight behavior of Canada geese. Auk 86, 671-681.
Ren, Q.M., Luo, S., Du, X.J., Chen, G.L., Song, S., Du, B., 2016. Helper effects in the azure-winged magpie Cyanopica cyana in relation to highly-clumped nesting pattern and high frequency of conspecific nest-raiding. J. Avian Biol. 47, 449-456.
Rendall, D., Notman, H., Owren, M.J., 2009. Asymmetries in the individual distinctiveness and maternal recognition of infant contact calls and distress screams in baboons. J. Acoust. Soc. Am. 125, 1792-1805.
Ripley, B., Venables, B., Bates, D.M., Hornik, K., Gebhardt, A., Firth, D., et al., 2013. Package 'mass'. .
Romani, F., Ramella Levis, E., Posillico, M., Opramolla, G., Pavan, G., 2022. Vocal repertoire of the Eurasian griffon vulture (Gyps fulvus) in the central Apennines: a baseline assessment. Bioacoustics 31, 283-312.
Seyfarth, R.M., Cheney, D.L., 2010. Production, usage, and comprehension in animal vocalizations. Brain Lang. 115, 92-100.
Seyfarth, R.M., Cheney, D.L., Bergman, T., Fischer, J., Zuberbühler, K., Hammerschmidt, K., 2010. The central importance of information in studies of animal communication. Anim. Behav. 80, 3-8.
Smith, C.L., 2017. Referential signalling in birds: the past, present and future. Anim. Behav. 124, 315-323.
Struhsaker, T.T., 1967. Auditory communication among vervet monkeys (Cercopithecus aetthiops). In: Altmann, S.A. (Ed.), Social Communication Among Primates. University of Chicago Press, Chicago, pp. 281-324.
Templeton, C.N., Greene, E., Davis, K., 2005. Allometry of alarm calls: Black-capped chickadees encode information about predator size. Science 308, 1934-1937.
Valencia, J., de la Cruz, C., González, B., 2003. Flexible helping behaviour in the azure-winged magpie. Ethology 109, 545-558.
Wang, L., Luo, Y., Maierdiyali, A., Chang, H., Ullah, S., Li, Z., 2021. Azure-winged Magpies Cyanopica cyanus passed the tasks on the Uzgiris-Hunt scale of object permanence. J. Ornithol. 162, 605-613.
Wang, L., Luo, Y., Wang, X., Maierdiyali, A., Chang, H., Li, Z., 2019. Azure-winged magpies solve string-pulling tasks by partial understanding of the physical cognition. Curr. Zool. 65, 385-392.
Wheeler, B.C., 2010. Production and perception of situationally variable alarm calls in wild tufted capuchin monkeys (Cebus apella nigritus). Behav. Ecol. Sociobiol. 64, 989-1000.
Wheeler, B.C., Fischer, J., 2012. Functionally referential signals: a promising paradigm whose time has passed. Evol. Anthropol. 21, 195-205.
Yambem, S.D., Chorol, S., Jain, M., 2021. More than just babble: functional and structural complexity of vocalizations of Jungle Babbler. Behav. Ecol. Sociobiol. 75, 118.
Table
2.
Estimated regression parameters, standard errors, profile confidence intervals (2.5% PCL and 97.5% PCL), χ2 values, df, and P values for each predictor variable to investigate the effects of season/year and sex (as fixed effects) on the migration distance, migration duration and migration timing of the Pied Avocet (Recurvirostra avosetta).


 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds