The Key Laboratory of Animal Resource and Epidemic Disease Prevention in Liaoning Province, School of Life Sciences, Liaoning University, Shenyang, 110036, China
Personality widely exists in diverse animal taxa. Such inter-individual variance in behaviour is supposed to be influenced by social context. However, it remains unknown whether the experience of social life has any carryover effects on the subsequent expression of personality. Here, we examined exploratory behaviour in caged Java Sparrows (Lonchura oryzivora) using exploration assays. Birds were assigned to live in either a solitary or a social context for four weeks. We compared the expressions of exploration before and after the treatments, and found that birds showed higher exploration tendencies after than before social life, while the isolated birds were consistent in their exploratory behaviours. Different living experience led to differences in the exploration activities for birds without significant differences in exploration before. Our results indicate that social experience can make birds more proactive.
Animal personality, as a series of consistent variances in behaviour among conspecifics across time and context, has been widely found in diverse taxa (Sih et al., 2004; Réale et al., 2007, 2010). The inter-individual variations of personality influence the way animals adapt to environment (Wolf and Weissing, 2012), with important ecological consequences (Sih et al., 2012; Wolf and Weissing, 2012). For example, high-exploratory Zebra Finches (Taeniopygia guttata) are less successful in finding food but more likely to adjust their behaviours (David et al., 2011). Since behaviour can be infinitely flexible in theory, how animals exhibit personality becomes a hot topic in behavioural ecology. Animal personality is considered as an adaptive outcome of inter-individual variance in state (Sih et al., 2015). The state of an individual can mean any features affecting the costs and benefits of its behavioural actions (Houston and McNamara, 1999), which includes features of the focal individual and characteristics of its ecological and social environment (Sih et al., 2015).
Social context could modulate individual behaviour (Webster and Ward, 2011; van den Bos et al., 2013). A growing body of evidence shows that the expression of personality depends on whether individuals are on their own or in the presence of companions (Webster and Ward, 2011). For instance, Java Sparrows (Lonchura oryzivora) are more active in the novel environment and the novel object tests in the presence of social partners (Zhang et al., 2020). In contrast, in a foraging experiment, Zebra Finches are bolder when isolated than in a flock (Kerman et al., 2018). Such modulating effects on animal personality vary across species and contexts, with conformity, facilitation, and inhibition being reported (Webster and Ward, 2011). Besides differences traced across genotypes, high plasticity of brain circuits in early phases of life (e.g., Jumping Spiders Marpissa muscosa, Liedtke et al., 2015; Black Widow Spiders Latrodectus Hesperus, DiRienzo et al., 2019), parts of which seem to retain their plasticity into adulthood, permits behavioural profiles to be shaped by environment during later phases of life (Gross and Hen, 2004; Champagne and Curley, 2005; Sachser et al., 2013).
While most studies have focused on the real-time effects of social context, empirical evidence on the carryover effects of social experience is relatively rare. Previous social experience in early life has been shown to affect the subsequent behaviour of fish, in that Cichlid Fish (Pelvicachromis taeniatus) reared in 90-day isolation are more aggressive and less willing to shoal than group-reared ones (Hesse and Thünken, 2014). It has been suggested that the more experience an individual has with a particular behaviour, the better it performs that behaviour (Sih et al., 2015). Since social context has been proved to greatly influence adult individuals, here comes the question how social experience would make the subsequent expression of personality be when individuals are alone once again. Jolles et al. (2016) showed that the social conditions two days before tests could obfuscate behaviour of Three-spined Sticklebacks (Gasterosteus aculeatus) in later contexts. While cumulative evidence has been found in fishes (e.g., Zebrafish Danio rerio, Kareklas et al., 2018; Three-spined Sticklebacks, Jolles et al., 2019; Munson et al., 2021), we still know little about the carryover effects of social experience on behaviour of animals from higher taxa, including birds.
The aim of our study was to investigate how social experience would affect the subsequent expression of personality in Java Sparrows. We randomly arranged birds for a four-week life in solitary or social treatment. Using a series of exploration assays, birds' exploratory behaviours were measured before and after the treatment. As birds might be influenced by conspecifics while in social treatment, we predicted that the social birds might behave differently from their behaviour before the treatment, whereas the solitary birds would be stable in exploration.
2.
Methods
2.1
Ethical note
All procedures involving birds were carried out in accordance with the Policy on the Care and Use of Animals, approved by the Ethical Committee, Center of Zoological Evolution and Systematic Zoological Museum of China, School of Life Sciences, Liaoning University (EC-LNU, 20200150). We adhered to the ASAB/ABS Guidelines for the use of animals. Birds' health was checked every day and no injuries were observed.
2.2
Study species and housing
Forty Java Sparrows (twenty males and twenty females) were purchased from a registered pet shop in Shenyang, Liaoning Province, China in September 2021. All birds were raised in farms in Dalian, Liaoning Province, and kept in family groups. Once the birds were fledged, they were transported to pet shops and housed singly (in cages measuring 25 × 25 × 25 cm). Subjects for this study were not related with each other and were transported to laboratory at the age of six months, without any prior breeding experience. Birds sexing was based on the sexual dimorphism of Java Sparrows (Soma and Iwama, 2017; Zhang et al., 2020). The birds were housed individually (in cages measuring 30 cm × 25 cm and 35 cm high) in the same laboratory for a month prior to the experiments. Both visual and auditory contacts were allowed, but physical contact was forbidden to ensure no mating. Each cage was equipped with a feeder, a drinker, a perch, and a nest. The birds were provided ad libitum with millets, health sand, fresh fruits, and water. The laboratory had a 12:12 h light cycle and the temperature was maintained at 22–24 ℃.
2.3
Experimental procedure
After a month of acclimatization in the laboratory, the exploration assays were proceeded twice for all birds with a three-week interval. To verify that the exploratory behaviour can be classified as a personality trait, behavioural consistency was assessed between the two-time trials for exploration tendencies (Réale et al., 2007). Once the second assays were completed, birds were randomly assigned to one of the two treatments: solitary (N = 20) or social (N = 20). Each treatment lasted for four weeks. Solitary birds were individually housed in 24-L cages (30 cm × 20 cm and 40 cm high), whereas the social birds were housed (10 per cage, males in one cage and females in another) in a 240-L cage (100 cm × 60 cm and 40 cm high) (Ros-Simó and Valverde, 2012; Appendix Fig. S1). Such arrangement aimed to maintain similar densities in both treatments (Munson et al., 2021). All cages were separated by opaque partitions to prevent birds from seeing conspecifics in adjacent compartments but acoustic contact was allowed (Apfelbeck and Raess, 2008; King et al., 2015). Enough food and water were provided in both treatments, with one perch and one nest per bird. After the 4-week treatment, all birds' explorations were tested individually for the third time.
2.4
Exploration assays
A novel environment test and a novel object test were involved in the exploration assays (Mainwaring et al., 2011). To limit neophobia involving boldness in the novel object test (Greggor et al., 2015), we used objects with which birds did not have any dangerous experience, in a non-threatening situation. Both tests were conducted during 8:00 to 16:00 on each test day. Birds initially encountered the novel environment test on the first day, and then the novel object test on the next two days (Mainwaring et al., 2011). The exploratory behaviour of each bird was recorded by a HP F860 driving recorder mounted on the top of the cage.
2.5
Novel environment test
On the first test day, exploratory behaviour in a novel environment was tested in an unfamiliar test cage (60 cm × 43 cm and 40 cm high; Appendix Fig. S2A). Five empty feeders were placed on the floor of the cage, with four at the corners and one in the middle. The focal bird was initially placed in a separated introductory cage, waiting 30 s for the sliding door to open, and could enter the test cage through the door spontaneously (Mainwaring et al., 2011). The whole test lasted for 10 min, and the time that each bird required to visit 4 of the 5 feeders was recorded by an observer (Q.C.) from the video. The time used by each bird was converted to a linear scale of 0–10 (0:00–0:59 = 10, 1:00–1:59 = 9, etc.; Martins et al., 2007). A score of 10 meant that the focal bird reached 4 of the 5 feeders within 1 min, while a score of 0 meant that the focal bird did not reach 4 different feeders within 10 min.
2.6
Novel object test
The novel object test was carried out on the next two test days to evaluate the response of birds to an unfamiliar object placed in the same test cage (Appendix Fig. S2B). The test cage was divided into three parts by two perches with the same size. Two novel objects (a battery measuring 4.5 by 1.5 cm and a carrot toy measuring 13 by 7 cm and 3 cm high; Appendix Fig. S3) were successively fixed on one of the two perches at random (Jha and Kumar, 2017). Birds were introduced to the battery on the second test day and the carrot toy on the third test day (Mainwaring et al., 2011). After 30-s waiting in the separated cage, the focal bird's behaviour was recorded by the driving recorder once upon entry. Each test lasted for 2 min. Following Jha and Kumar (2017), the reactions to both objects were recorded on a scale of 0–5 (0 meant to stay on the floor furthest from the novel object; 1 meant to stay on the floor between the two perches; 2 meant to stay on the floor but be close to the perch with the novel object; 3 meant to hop on the empty perch; 4 meant to hop on the perch with the novel object but not touch it; and 5 meant to hop on the perch with the novel object and touch it). The score was the maximum rating achieved when the bird presented more than one of the five situations above (Mainwaring et al., 2011).
The exploration score of each bird was calculated as the sum of their performances recorded in the novel environment test and the novel object test (Drent et al., 2003; van Oers et al., 2004). In theory, the lowest score is 0 and the highest score is 20. Actually, few birds reached four empty feeders in the 10-min novel environment test, with only one bird getting a score in the first-time test (similar phenomenon in Java Sparrows see Wang et al., 2022). Thus, the maximum pretreatment score recorded was 10 (Martins et al., 2007).
2.7
Statistical analysis
We tested the repeatability of all forty birds' explorations across the twice repeated behavioural assays before treatments by calculating average measures intraclass correlation coefficients (ICCs) and 95% confidence intervals (CIs) with two-way random effects model, using SPSS v. 21.0 (IBM, Armonk, NY, U.S.A.).
To assess whether different social experience could affect birds' exploration, a linear mixed model (LMM) was fitted in the 'nlme' package of R (R Development Core Team, 2019). Treatment (solitary, social), stage (pre, post), sex (male, female), the interaction treatment * stage and the interaction treatment * sex were included as the explanatory variables in our model, while the exploration score (average of the first two replicates as the pretreatment score) was included as the dependent variable. We fitted bird ID as a random factor in model to control for individual differences and repeated observations. Model fit was checked by visual inspection of quantile-quantile plots of model residuals versus the predictor. Differences in exploration scores were compared among the groups in the same stage and across stages, using independent sample t-tests or paired t-tests. Relationship between pre- and post-treatment exploration scores of social birds was investigated using Pearson correlations.
3.
Results
A significant proportion of observed variation across all the forty birds in the two repeated behavioural assays before treatments could be attributed to inter-individual differences (ICC (CI) = 0.551 (0.295–0.734), F39, 39 = 3.659, P < 0.001).
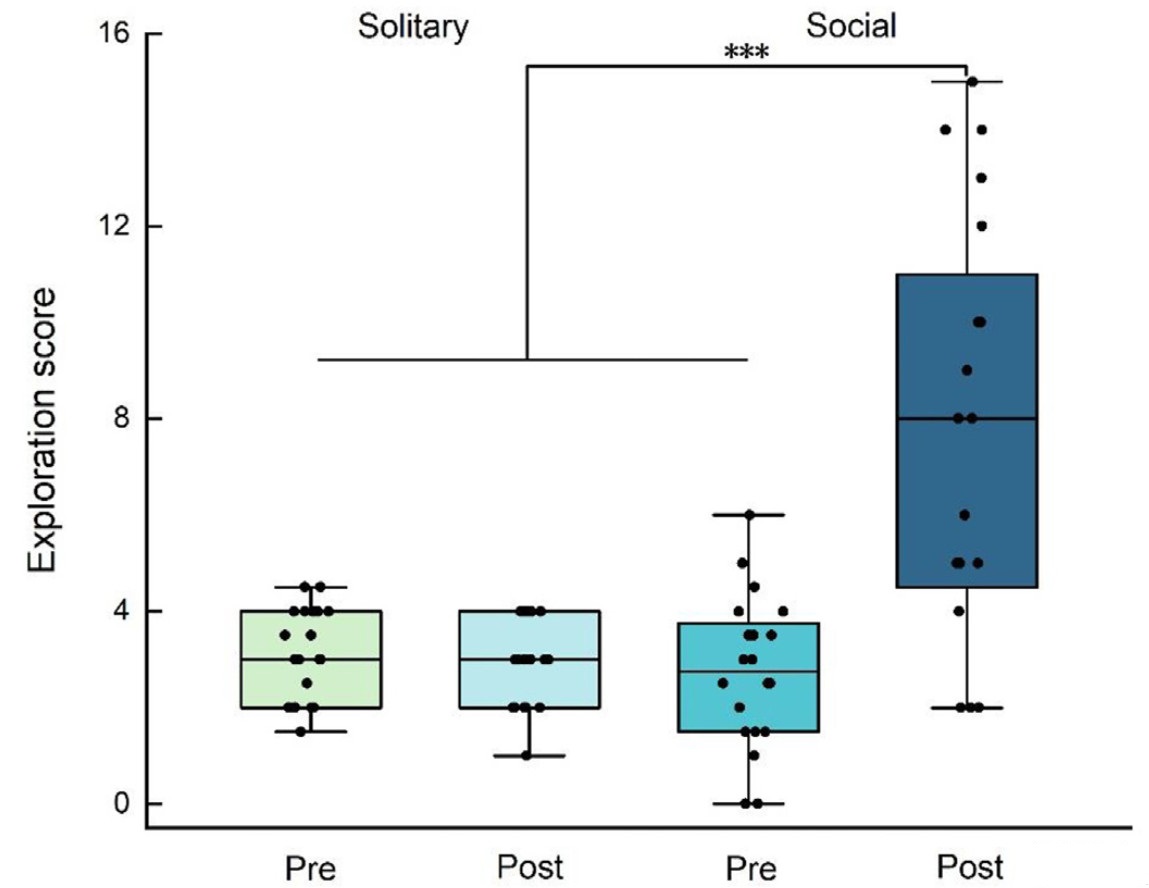
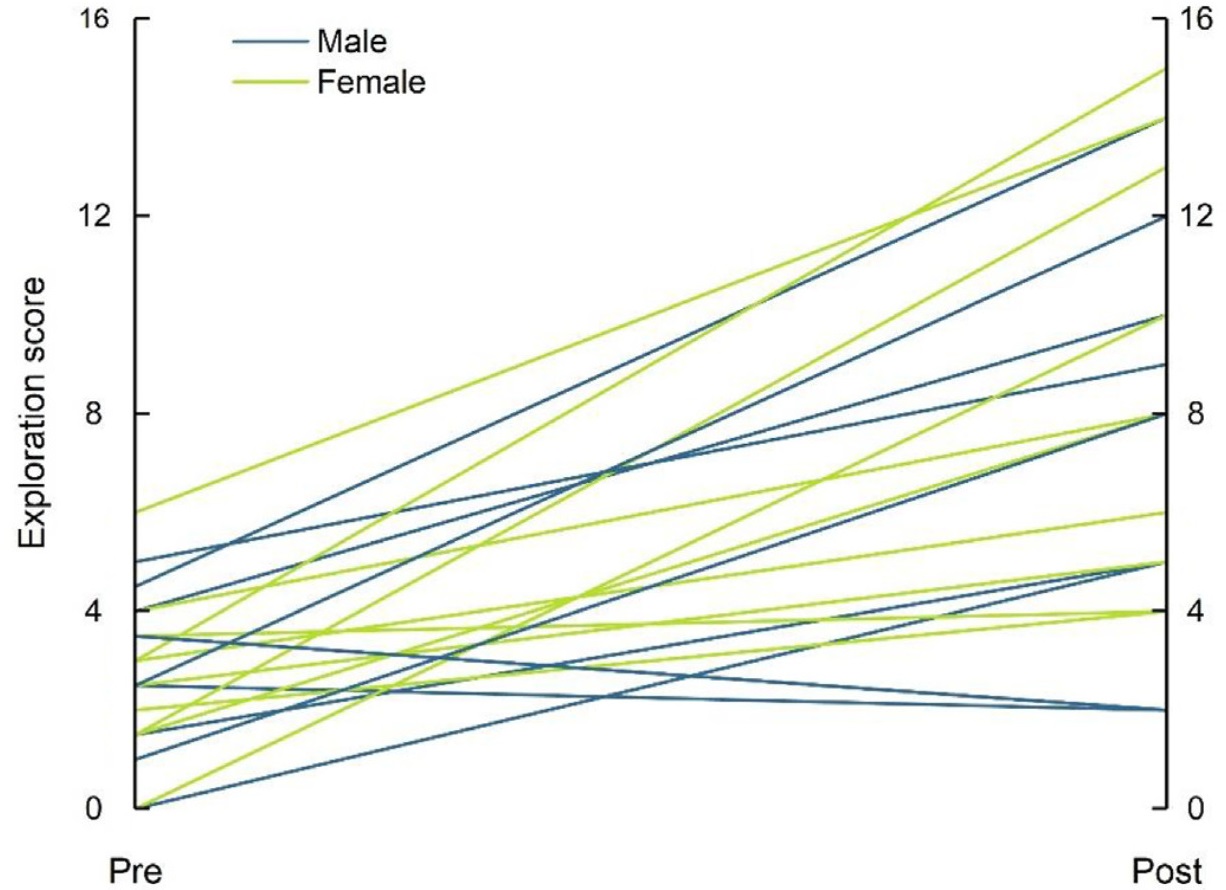
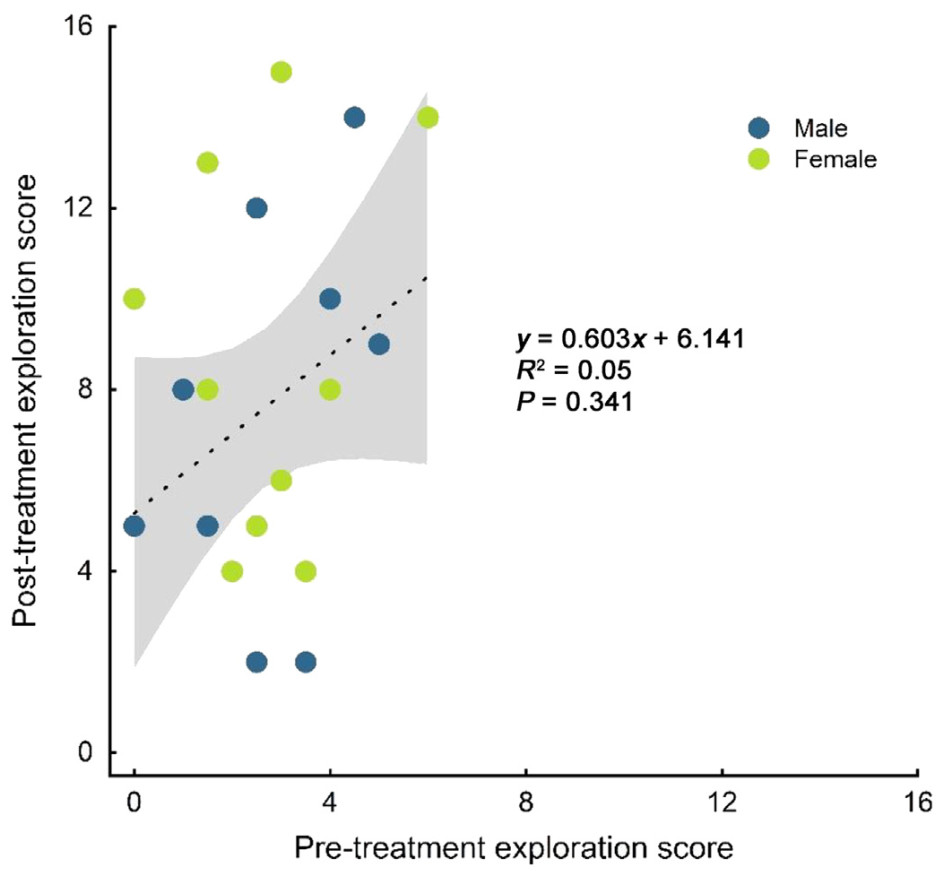
We found an interaction between treatment and stage on birds' exploration scores (LMM: estimate ± SE = −5.200 ± 0.720, t = −7.221, P < 0.001; Table 1, Fig. 1). Exploration scores showed no variation between males and females (LMM: estimate ± SE = −0.533 ± 0.685, t = −0.779, P = 0.441; Table 1, Fig. 1), and both sexes showed similar reactions to the treatments (LMM: estimate ± SE = 0.500 ± 0.969, t = 0.516, P = 0.609; Table 1, Fig. 1). For the two treatments, birds showed no significant difference in pretreatment scores (P = 0.401; Fig. 1), but in post-treatment scores (P < 0.001; Fig. 1). Solitary birds did not differ in their pre- and post-treatment exploration scores (t = 0.809, P = 0.428; Fig. 1), whereas social birds showed significantly higher exploration scores after treatment than before (t = −5.407, P < 0.001; Fig. 1), with 18 out of the 20 individuals showing higher exploration scores (Fig. 2). There was no significant correlation between the pre- and post-treatment exploration scores of social birds (rs = 0.224, N = 20, P = 0.341; Fig. 3).
Table
1.
Results from linear mixed model (LMM) testing the effects of treatment, stage, sex, and their interactions on birds' exploration scores.
Figure
1.
The exploration scores of birds pre- and post-solitary or social treatment. Box plots show 25th, 50th (median), and 75th percentiles with horizontal lines. The whiskers end at the largest and smallest non-outliers. ***p < 0.001.
In this study, we tested whether the experience of social life could affect the subsequent expression of personality. We found that social experience made birds differ in their pre- and post-treatment explorations. While the solitary birds maintained consistent exploratory behaviours, social birds showed higher exploration tendencies after the four-week social life than before. However, the magnitude of increases in the social birds' exploration scores were different, which had no correlation with the starting condition.
We did not find behavioural consistency across contexts as almost all birds scored 0 in the repeated novel environment tests before treatments. Similar phenomena were observed in Zebra Finches (Martins et al., 2007). The increase in post-treatment scores came from the birds' getting scores in the novel environment test and their higher scoring in the novel object test. However, there was no significant consistency between the post-treatment scores in the novel environment test and the novel object test (ICC (CI) = −0.307 (−0.653–0.145), F39, 39 = 0.530, P = 0.912). It is unclear whether such inconsistency is due to the influence of social experience or the possible different responses of individuals across exploration tasks, as proposed by Miller et al. (2022).
Our results support the prediction that social experience would have a carryover effect on the subsequent expression of personality in Java Sparrows, which is in line with previous findings (Hsu and Wolf, 1999; Frost et al., 2007; Jolles et al., 2014). We interpreted it as that the solitary birds almost had no need to adjust and conform their behaviour to maintain or reach coherence in any population (e.g., Ioannou and Dall, 2016). They remained their prior expressions of exploration just because they received no impact from conspecifics. There was a general increase in the exploration tendencies of twenty birds with social experience. Social context can offer animals more safety than solitude (Magurran and Pitcher, 1983; Webster and Ward, 2011; Ward, 2012), and thus risk-aversive behaviour may be less facilitated in identical situations (Kareklas et al., 2018). Moreover, resource competition among conspecific individuals could also lead to a higher dispersal tendency (Hauzy et al., 2007). Even when tested alone, the expression of individual's personality could still be affected (Jolles et al., 2016), probably because social experience can carry over from one context to the next (Frost et al., 2007; Gómez-Laplaza, 2009; Jolles et al., 2014).
Although social experience indeed promoted the general proactive behaviour in two flocks, the uncorrelated relationship between the birds' pre- and post-treatment exploration scores for the social treatment suggested that individual's future expression of personality could not be simply predicted by its initial level. As a key component of dispersal to habitat with higher quality (Bowler and Benton, 2005; Debeffe et al., 2013), exploration would be a means for the individual to obtain fitness benefits limited to the individual's current rank. It is notable that there is a trade-off between such fitness benefits and the potential cost of dispersal (Coates et al., 2019). Despite in the same social context, birds with different personalities may still encounter selective pressures to different extents (Greenberg and Mettke-Hofmann, 2001; Réale et al., 2007). Those variances in pressures probably serve as an explanation for the various ways through which birds adjust their behaviours. Further works should consider the rank of each individual. It is notable that there were significantly higher post-treatment exploration scores of the birds in the social treatment rather than the solitary one. As high exploration could help individuals expand their territories and obtain more resources, such variation might bring advantages to the population.
In summary, our results indicate that social experience can have a carryover effect on the subsequent expression of personality. We examined such effect respectively in male and female populations, because we have no means to deal with the influence of potential breeding in a mixed group. For the birds with social experience, they showed higher exploration tendencies in general than themselves before. Such social experience could lead to a more proactive average expression of birds flocking than the birds which were not different from the flocks initially but lived alone. Nevertheless, the bird flocking's initial exploration tendencies did not predict the increase in their exploratory behaviours after the social experience. For the need of rearing, we did not control the social living experience in their family groups before the experiment. The same density kept, that in fact allowed the social birds to have much more to explore, might also promote the birds' exploratory behaviour. Further works should be done to address these issues. To understand more about the carryover effects of social experience, we call for more experiments on various group compositions and varying durations of the social experience. The recovery of individual's behavioural expressions may also deserve more attention.
Ethics statement
The ethics standard is provided in the section "2.1. Ethical note" in the main text.
Authors' contributions
JY and QC designed the experiment. GC, MS, YW and QC collected the data. JW and QC conducted the analyses. QC and JY wrote the manuscript. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Authors thank two anonymous reviewers for their constructive comments and suggestions. This work was supported by Department of Science and Technology of Liaoning Province (2019-ZD-0196) and Department of Education of Liaoning Province (LJC202009). All authors declare no conflict of interests.
Apfelbeck, B., Raess, A.M., 2008. Behavioural and hormonal effects of social isolation and neophobia in a gregarious bird species, the European starling (Sturnus vulgaris). Horm. Behav. 54, 435-441. .
Bowler, D.E., Benton, T.G., 2005. Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biol. Rev. 80, 205-225. .
Champagne, F.A., Curley, J.P., 2005. How social experiences influence the brain. Curr. Opin. Neurobiol. 15, 704-709. .
Coates, W.D., Hale, R., Morrongiello, J.R., 2019. Dispersal decisions and personality in a freshwater fish. Anim. Behav. 157, 209-218. .
David, M., Cezilly, F., Giraldeau, L.A., 2011. Personality affects zebra finch feeding success in a producer-scrounger game. Anim. Behav. 82, 61-67. .
Debeffe, L., Morellet, N., Cargnelutti, B., Lourtet, B., Coulon, A., Gaillard, J. M., et al., 2013. Exploration as a key component of natal dispersal: dispersers explore more than philopatric individuals in roe deer. Anim. Behav. 86, 143-151. .
DiRienzo, N., Johnson, J.C., Dornhaus, A., 2019. Juvenile social experience generates differences in behavioral variation but not averages. Behav. Ecol. 2, 455-464. .
Drent, P., van Oers, K., van Noordwijk, A.J., 2003. Realized heritability of personalities in the great tit (Parus major). Philos. T. Roy. Soc. B 270, 45-51. .
Frost, A.J., Winrow-Giffen, A., Ashley, P.J., Sneddon, L.U., 2007. Plasticity in animal personality traits: does prior experience alter the degree of boldness? Philos. T. Roy. Soc. B 274, 333-339. .
Gómez-Laplaza, L.M., 2009. Recent social environment affects colour-assortative shoaling in juvenile angelfish (Pterophyllum scalare). Behav. Process. 82, 39-44. .
Greenberg, R., Mettke-Hofmann, C., 2001. Ecological aspects of neophobia and neophilia in birds. Curr. Ornithol. 16, 119-178. .
Greggor, A.L., Thornton, A., Clayton, N.S., 2015. Neophobia is not only avoidance: improving neophobia tests by combining cognition and ecology. Curr. Opin. Behav. Sci. 6, 82-89. .
Gross, C., Hen, R., 2004. The developmental origins of anxiety. Nat. Rev. Neurosci. 5, 545-552. .
Hauzy, C., Hulot, F.D., Gins, A., Loreau, M., 2007. Intra-and interspecific density-dependent dispersal in an aquatic prey-predator system. J. Anim. Ecol. 76, 552-558. .
Hesse, S., Thünken, T., 2014. Growth and social behavior in a cichlid fish are affected by social rearing environment and kinship. Naturwissenschaften 101, 273-283. .
Houston, A.I., McNamara, J.M., 1999. Models of Adaptive Behaviour. Cambridge University Press, London.
Hsu, Y., Wolf, L., 1999. The winner and loser effect: integrating multiple experiences. Anim. Behav. 57, 903-910. .
Ioannou, C.C., Dall, S.R.X., 2016. Individuals that are consistent in risk-taking benefit during collective foraging. Sci. Rep. 6, 33991. .
Jha, N.A., Kumar, V., 2017. Effect of no-night light environment on behaviour, learning performance and personality in zebra finches. Anim. Behav. 132, 29-47. .
Jolles, J.W., Fleetwood-Wilson, A., Nakayama, S., Stumpe, M.C., Johnstone, R.A., Manica, A., 2014. The role of previous social experience on risk-taking and leadership in three-spined sticklebacks. Behav. Ecol. 25, 1395-1401. .
Jolles, J.W., Taylor, B.A., Manica, A., 2016. Recent social conditions affect boldness repeatability in individual sticklebacks. Anim. Behav. 112, 139-145. .
Jolles, J.W., Briggs, H., Araya-Ajoy, Y.G., Boogert, N.J., 2019. Personality, plasticity and predictability in sticklebacks: bold fish are less plastic and more predictable than shy fish. Anim. Behav. 154, 193-202. .
Kareklas, K., Elwood, R.W., Holland, R.A., 2018. Grouping promotes risk-taking in unfamiliar settings. Behav. Process. 148, 41-45. .
Kerman, K., Miller, L., Sewall, K., 2018. The effect of social context on measures of boldness: zebra finches (Taeniopygia guttata) are bolder when housed individually. Behav. Process. 157, 18-23. .
King, A.J., Williams, L.J., Mettke-Hofmann, C., 2015. The effects of social conformity on Gouldian finch personality. Anim. Behav. 99, 25-31. .
Liedtke, J., Redekop, D., Schneider, J.M., Schuett, W., 2015. Early environmental conditions shape personality types in a jumping spider. Front. Ecol. Evol. 3, 134. .
Magurran, A.E., Pitcher, T.J., 1983. Foraging, timidity and shoal size in minnows and goldfish. Behav. Ecol. Sociobiol. 12, 147-152. .
Mainwaring, M.C., Beal, J.L., Hartley, I.R., 2011. Zebra finches are bolder in an asocial, rather than social, context. Behav. Process. 87, 171-175. .
Martins, T., Roberts, M., Giblin, I., Huxham, R., Evans, M., 2007. Speed of exploration and risk-taking behavior are linked to corticosterone titres in zebra finches. Horm. Behav. 52, 445-453. .
Miller, R., Lambert, M.L., Frohnwieser, A., Brecht, K.F., Bugnyar, T., Crampton, I., et al., 2021. Socio-ecological correlates of neophobia in corvids. Curr. Biol. 32, 74-85. .
Munson, A., Michelangeli, M., Sih, A., 2021. Stable social groups foster conformity and among-group differences. Anim. Behav. 174, 197-206. .
Réale, D., Reader, S.M., Sol, D., Mcdougall, P.T., Dingemanse, N.J., 2007. Integrating animal temperament within ecology and evolution. Biol. Rev. 82, 291-318. .
Réale, D., Dingemanse, N.J., Kazem, A., Wright, J., 2010. Evolutionary and ecological approaches to the study of personality. Philos. T. Roy. Soc. B 365, 3937-3946. .
Ros-Simó, C., Valverde, O., 2012. Early-life social experiences in mice affect emotional behaviour and hypothalamic-pituitary-adrenal axis function. Pharmacol. Biochem. Behav. 102, 434-441. .
Sachser, N., Kaiser, S., Hennessy, M.B., 2013. Behavioural profiles are shaped by social experience: when, how and why. Philos. T. Roy. Soc. B 368, 20120344. .
Sih, A., Bell, A.M., Johnson, J.C., 2004. Behavioral syndromes: an ecological and evolutionary overview. Trends Ecol. Evol. 19, 372-378. .
Sih, A., Cote, J., Evans, M., Fogarty, S., Pruitt, J., 2012. Ecological implications of behavioural syndromes. Ecol. Lett. 15, 278-289. .
Sih, A., Mathot, K.J., Moirón, M., Montiglio, P.O., Wolf, M., Dingemanse, N.J., 2015. Animal personality and state-behaviour feedbacks: a review and guide for empiricists. Trends Ecol. Evol. 30, 50-60. .
Soma, M., Iwama, M., 2017. Mating success follows duet dancing in the Java Sparrow. PLoS One 12, e0172655. .
van den Bos, R., Jolles, J.W., Homberg, J.R., 2013. Social modulation of decision-making: a cross-species review. Front. Hum. Neurosci. 7, 301. .
van Oers, K., Drent, P.J., De Goede, P., 2004. Realized heritability and repeatability of risk-taking behaviour in relation to avian personalities. Philos. T. Roy. Soc. B 271, 65-73. .
Ward, A.J.W., 2012. Social facilitation of exploration in mosquitofish (Gambusia holbrooki). Behav. Ecol. Sociobiol. 66, 223-230. .
Wang, J.Y., Wang, D.P., Chen, Q.Y., Zhang, J., Racey, P., Jiang, Y.T., et al., 2022. Female Java Sparrows prefer high exploratory males without assortative mating. Behav. Process. 200, 104671. .
Webster, M.M., Ward, A., 2011. Personality and social context. Biol. Rev. Camb. Philos. Soc. 86, 759-773. .
Wolf, M., Weissing, F.J., 2012. Animal personalities: consequences for ecology and evolution. Trends Ecol. Evol. 27, 452-461. .
Zhang, J., King, A.J., Fürtbauer, I., Wang, Y.W., He, Y.Q., Zhang, Z.W., et al., 2020. Facilitative effects of social partners on Java Sparrow activity. Anim. Behav. 161, 33-38. .

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds