The effect of urbanization and exposure to multiple environmental factors on life-history traits and breeding success of Barn Swallows (Hirundo rustica) across China
State Key Laboratory of Biocontrol, School of Ecology, Sun Yat-sen University, Guangzhou, 510006, China
b.
Institute of Eco-Environmental Research, Guangxi Academy of Sciences, Nanning, 530007, China
c.
Department of Health and Environmental Sciences, Xi'an Jiaotong-Liverpool University, Suzhou, 215028, China
d.
Key Laboratory for Biodiversity Science and Ecological Engineering, Ministry of Education, College of Life Sciences, Beijing Normal University, Beijing, 100091, China
e.
College of Wildlife and Protected Area, Northeast Forestry University, Harbin, 150006, China
f.
Institute of Wildlife Conservation, Central South University of Forestry and Technology, Changsha, 410004, China
g.
College of Life Science, Liaoning University, Shenyang, 110036, China
h.
Department of Ecology and Evolutionary Biology, University of Colorado, Boulder, USA
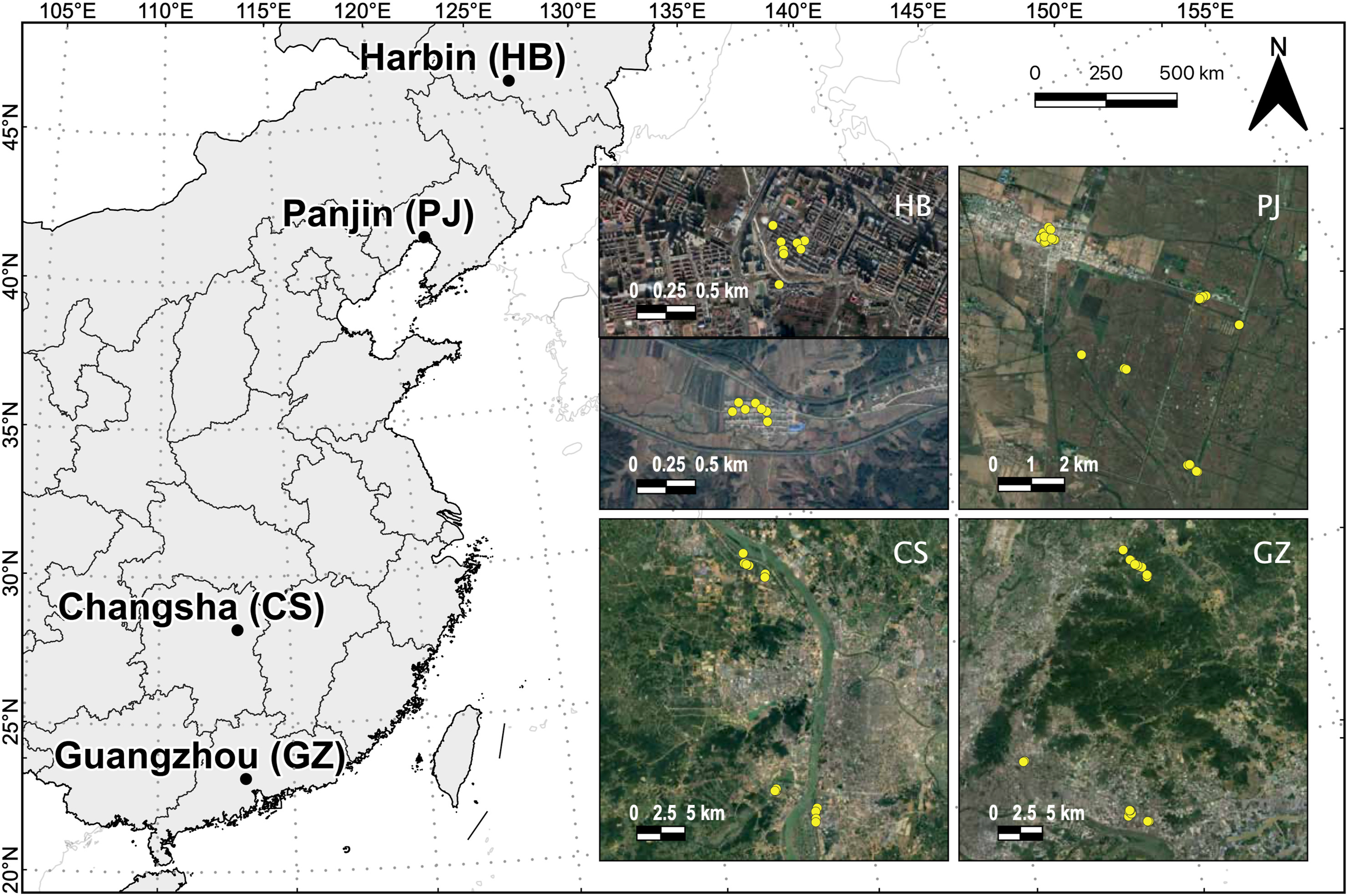
In addition to landscape changes, urbanization also brings about changes in environmental factors that can affect wildlife. Despite the common referral in the published literature to multiple environmental factors such as light and noise pollution, there is a gap in knowledge about their combined impact. We developed a multidimensional environmental framework to assess the effect of urbanization and multiple environmental factors (light, noise, and temperature) on life-history traits and breeding success of Barn Swallows (Hirundo rustica) across rural to urban gradients in four locations spanning over 2500 km from North to South China. Over a single breeding season, we measured these environmental factors nearby nests and quantified landscape urbanization over a 1 km2 radius. We then analysed the relationships between these multiple environmental factors through a principal component analysis and conducted spatially explicit linear-mixed effects models to assess their effect on life-history traits and breeding success. We were particularly interested in understanding whether and how Barn Swallows were able to adapt to such environmental conditions associated with urbanization. The results show that there is significant variation in the exposure to environmental conditions experienced by Barn Swallows breeding across urbanization gradients in China. These changes and their effects are complex due to the behavioural responses ameliorating potential negative effects by selecting nesting sites that minimize exposure to environmental factors. However, significant relationships between landscape urbanization, exposure to environmental factors, and life-history traits such as laying date and clutch size were pervasive. Still, the impact on breeding success was, at least in our sample, negligible, suggesting that Barn Swallows are extremely adaptable to a wide range of environmental features.
In order to coordinate the behaviour and activities with conspecifics, various signals are needed for animals to communicate with each other (Manser, 2016). Vocal communication is commonly used in mammals and birds. Unlike human language, animal vocalisations largely depend on external objects and events, such as the presence of predators or food (Macedonia and Evans, 1993). According to previous studies, many animals would use specific call types in similar contexts, and the connection between call type and context could hardly be separated (Seyfarth and Cheney, 2010). Therefore, the current studies on animal vocalisations were mainly conducted under the framework of 'functional referential signals', which was first put forward by Macedonia and Evans (1993) and referred to the strong connection between the context (information content) of the signal and the response of the receiver (Wheeler and Fischer, 2012).
In early studies, however, the information content has been questioned by several researchers, arguing that the concept of information transfer should be abandoned in the study of animal vocalisations (Seyfarth et al., 2010). The motivational structural rules were proposed to explain the production of animal vocal communication (Morton, 1977). As found in various vertebrates, calls emitted under low aggressive motivation conditions would have a narrow bandwidth and high frequency, while those under high aggressive motivation would have a wide bandwidth and low frequency. The physical structures of calls were believed to have a general relationship with the motivation of the signaller. The relationship may significantly affect the evolution of animal vocalisations (Morton, 1977). Similarly, in the perception of animal vocal communication, the 'affect conditioning' model was put forward by Owren and Rendall (1997), highlighting the effect of the acoustic structure on the receiver. For example, the distress screams of primates could influence the nervous systems and automatic response processes of the receiver (Rendall et al., 2009).
Nevertheless, with increasing potential information being found, along with the changes in behaviour after acquiring such information, the information hypothesis was recognised to be of great importance in driving the field of study forward (Seyfarth et al., 2010). Multiple information processes have been corroborated to play an essential role in the production and perception of animal vocalisations, rather than the motivation or emotional status of the signaller merely (Seyfarth and Cheney, 2010). For instance, the Dwarf Mongooses (Helogale parvula) could give two predator-specific alarm calls to terrestrial and aerial predators respectively and induce different responses of conspecifics (run for shelter in response to an aerial alarm; scan the area horizontally in response to a terrestrial alarm; Beynon and Rasa, 1989). They would react the same way after receiving these alarm calls even if the context of the predator was not provided (Collier et al., 2017). The same call type emitted in different contexts could provide different information and induce different responses. Such information may be edited through the number of elements (Templeton et al., 2005), the sequence of the elements within the call (Fishbein et al., 2020) and even the subtle difference in the spectral parameters (Dutour et al., 2021).
In birds, calls are simple vocalisations and the note is the basic element of a call. Based on the number of notes within a call, bird vocalisations could be classified as monosyllabic or multisyllabic calls. Additionally, a more complex and ordered vocalisation, song, could be emitted by some species, mainly in the context of territory establishment, maintenance and mate attraction (Marler, 2008). Azure-winged Magpies (Cyanopica cyanus) belong to the Corvidae family, which tend to have inconspicuous songs but large repertoires of calls (Marler, 2008). They are cooperatively breeding birds and have been reported to show proactive prosociality (Komeda et al., 1987; Horn et al., 2016). The 'social complexity hypothesis' pointed out that the complexity of communication tends to be positively related to social complexity (Blumstein and Armitage, 1997). However, little attention was paid to their vocalisation, let alone the comparison of their communication system to other birds. By contrast, the breeding ecology and cognitive study of Azure-winged Magpies have been investigated deeply to date (Ren et al., 2016; Da et al., 2018; Wang et al., 2019; Massen et al., 2020; Zhang et al., 2021). Given that a complex communication system is of great importance for individuals living in organised groups to respond to conspecifics appropriately (Grieves et al., 2015; Boucherie et al., 2019), this imbalanced development may impede further investigation in Azure-winged Magpies. Therefore, a detailed study on the structure and function of vocal communication is indispensable.
Our study aimed to give a detailed description of the context and function of each call type of Azure-winged Magpies. The first aspect of this study was to examine the vocal repertoire of Azure-winged Magpies and classify the vocalisations based on their acoustic parameters. According to the 'social complexity hypothesis', we hypothesised that Azure-winged Magpies should have a complex communication system with a relatively large vocal repertoire. The second aspect was to investigate the association between vocalisation categories and contexts, based on which we could interpret their function. If the vocalisations are largely dependent on external events, then a strong connection with the surroundings is expected to be seen. Meanwhile, if there exist 'functional referential signals' in Azure-winged Magpies, then these call types should induce a particular response of the receiver.
2.
Methods
2.1
Study sites and species
The study was conducted on the free-ranging Azure-winged Magpies between September 2021 and September 2022 in Nanjing University, Nanjing, Jiangsu Province, China (32°7′12″ N, 118°57′11″ E). Five field locations were studied on campus and each location was precisely within the territory of one specific flock. The territory was measured according to the tracking and monitoring simultaneously conducted by multiple experimenters before the experiment and over the entire study period. The territory of each flock has no overlap with each other and no flock was spotted crossing its own territory to other field locations. Thus, we could identify the flocks according to the location without capturing and marking the individuals. The number of individuals within each flock ranges from 29 to 42 (36 on average), with the newborn birds not being accounted.
The flock had an average sex ratio of 1.3 males per female and was not significantly different from 1:1 according to the early study (Valencia et al., 2003). However, due to the barely discernible difference between sexes, we did not assign the sex composition. Azure-winged Magpies are cooperative breeders and have flexible helping behaviour, with nearly 50% of nests being assisted by helpers at nest (Valencia et al., 2003). The helpers are predominantly males and mainly contribute to the provision of the incubating female and the brood, as well as the defence of nest (Ren et al., 2016). The breeding season of Azure-winged Magpies in this area started from late April to late August.
2.2
Data collection
A recorder (TASCAM DR-05X), connected to a directional microphone (Sennheiser ME66/K6) was used to record all vocalisations (44.1 kHz sampling rate, 16 bits sample size, WAV format). Two approaches were used during the acoustic data collection: automatic and manual recordings. Automatic recordings were carried out by fixing the recorder and microphone on a 1.5 m tall metal pole under the nest during the breeding season (May–August 2022) and on the foraging sites during the non-breeding season (October 2021–March 2022). The recording took place from 7:00 a.m. to 7:00 p.m. on sunny days (i.e., a 12-h continuous recording). Manual recordings were carried out by tracking the target flock from 5:30 a.m. to 8:30 a.m. and 3:30 p.m. to 6:30 p.m. during both breeding and non-breeding seasons.
The context data were collected separately from the recording of vocalisations, using an 8 × 42 binocular (Vector Optics Forester, China) from 5:30 a.m. to 8:30 a.m. and 3:30 p.m. to 6:30 p.m. during both breeding and non-breeding seasons. By scanning immediately after we focused on the signaller, the category, location and behaviour of both signallers and receivers were noted, along with the surroundings. The category included: (1) nestlings; (2) juveniles; (3) adults. Juveniles and adults were distinguished according to plumage and the top of the head (Harada, 1997). Juveniles have white-mottled heads and duller plumage while adults have a glossy black top to the head and brighter plumage. Location included: (1) within nest; (2) around nest; (3) ground (away from nest); (4) on branch (away from nest); (5) in the air (away from nest). Behaviour included: (1) food intake (FI), when individuals are foraging or receiving food from others; (2) rest (R), when individuals were resting on the branch with no other behaviours; (3) play (P), when individuals were chasing with each other from one branch to another; (4) movement (M), individual change its position through short or long flight, being divided into approach the signaller (AS) and follow the signaller (FS) for behaviour of the receiver; (5) alert (A), alert to danger in the non-breeding season or not around the nest in the breeding season; (6) incubation or brooding (IB), when parents are maintaining the temperature of eggs or nestlings in the nest; (7) food provision (FP), when individuals are providing food for the young or incubating females; (8) beg for food (BF), individuals begging food from others; (9) copulation (C); (10) defence of nest (DN), adults defending the nest from any intruders or predators in the breeding season; (11) food competition (FC), when individuals competing food with each other, including fight, displacement and acoustic warnings; (12) individual alone (IA), when individuals were separated from the group, for behaviour of the signallers only. Surroundings included: (1) no hints of danger (ND), no danger was found around the signaller; (2) presence of intruder (PI), potential competitors or a threat around nests, normally Eurasian Magpies (Pica pica), Red-billed Blue Magpie (Urocissa erythrorhyncha) and Asian Barred Owlet (Glaucidium cuculoides); (3) presence of predator (PP), when a predator is identified such as a free-ranging Cat (Felis silvestris catus) or a Eurasian Sparrowhawk (Accipiter nisus). The behaviour of other individuals would be counted as a response only if there was a strong connection with the vocalisation of the signaller (i.e., when such a behavioural change was involved with the signaller and occurred immediately after the vocalisation). The next piece of data would not be recorded in 5 min to ensure that the context of one specific vocalisation is independent of each other. All the data were then used to interpret their potential functions, while the category and the location were not included in the statistical analysis to better focus on our objective.
2.3
Acoustic analysis
Raven Pro 1.6 (Cornell Laboratory of Ornithology, USA) was used to plot the spectrograms and conduct all acoustic analyses. The spectrograms were generated using Hann window type, DFT of 512 samples and an overlap of 50%, with 0.0058 s time resolution and 86.10 Hz frequency resolution. Elements were considered to belong to the same call if the interval was < 0.1s (Catchpole and Slater, 2008). In this way, the vocalisations were distinguished into monosyllabic (only one element) or multisyllabic (more than one element). Only calls of high quality were selected for further analysis. Once a call was selected, any other calls emitted within the next 5 min would not be considered to largely avoid the continuous record of vocalisations from the same signaller.
Eight parameters were extracted from these calls, including (a) spectral parameters: (1) frequency 5% (F5), the frequency that divides the spectrum into two intervals which contain 5% and 95% of the energy; (2) frequency 95% (F95), the frequency that divides the spectrum into two intervals which contain 95% and 5% of the energy; (3) peak frequency (PF), the frequency at which peak power occurs; (b) harmonic parameters: (4) F0min, the minimum frequency of the fundamental harmony; (5) F0max, the maximum frequency of the fundamental harmony; (6) Fdom, the mean frequency of the dominant harmony (the harmony with the greatest power); (c) temporal parameters: (7) duration (Dur), the duration of the first element of the vocalisation and (d): (8) Num, the total number of elements (for multisyllabic calls only).
For vocalisations with no harmonies, the harmonic parameters were standardised, with F0min representing the minimum frequency of the call, F0max being equal to the frequency 95% and Fdom being the average of F0min and F0max.
2.4
Statistical analysis
Twelve categories (seven monosyllabic calls and five multisyllabic calls) were defined according to the aural and visual details of both manual and automatic recordings. A discriminant function analysis (DFA; lda in R program using package MASS; Ripley et al., 2013) on the acoustic parameters of monosyllabic and multisyllabic calls was carried out respectively to validate the initial identification of call types. A leave-one-out cross-validation DFA was then performed in the same way and generated the percentage of correct classifications (CC). All parameters used for the analysis were log-transformed, considering that it is sensitive to outliers and the data should meet linearity assumptions (Romani et al., 2022).
The difference between the sound categories was tested by a non-parametric multivariate permutational analysis of variance (PERMANOVA; adonis2 in R program using package vegan; Oksanen et al., 2019). The Euclidean distance method was selected and 999 permutations were used. The variables used for PERMANOVA are the same as those used for DFA and log-transformed as well.
To reflect the general difference between the contexts of all call types, a Chi-square test was conducted on the frequency of twelve call types observed in different contexts. Given that the expected frequency values of some contexts were less than one, we grouped some similar contexts to meet the assumptions of the test: presence of intruder and presence of predator were mixed as presence of danger; food intake, rest and play were combined, as they were the basic activities of individuals which were induced by internal needs rather than external stimuli; incubation or brooding, feed, beg for food and copulation were combined, as they were all close interactions between the young and the adults or among the couples and helpers related to the reproductive behaviour; food competition was mixed with defence of nest, since they were both competitive behaviours. Therefore, a 12 × 2 contingency table was used for the analysis of surroundings and a 12 × 6 contingency table for behaviour of the signaller and behaviour of the receiver. To figure out the detailed difference of a specific call type between contexts, a Chi-square goodness-of-fit test was applied to each call type respectively. These analyses were performed using R software version 4.2.1 (R Core Team, 2022).
3.
Results
Twelve distinct call types were assigned in this study. All calls and the context of call types were recorded in each flock (Appendix Table S1). The category of calls had a strong correlation with the contexts and each specific call type was dependent on different contexts as well (Appendix Table S2).
3.1
Statistical classification of call types
A total of 357 vocalisations (230 for monosyllabic calls and 127 for multisyllabic calls) were extracted from the 95 h of recordings (63 h from manual recordings and 32 h from automatic recordings). Based on the aural and visual details of the recordings, the vocalisations were grouped into 12 categories (7 monosyllabic calls and 5 multisyllabic calls) preliminarily and were named from 'A' to 'L' according to the initial order of these vocalisations recorded by the experimenter. The parameters used for linear discriminant analysis and PERMANOVA are shown in Table 1.
Table
1.
Acoustic parameters of twelve vocalisation categories of Azure-winged Magpies showing mean ± SE.
Category
n
F0min
F0max
Fdom
F5
F95
PF
Dur
Num
A
27
2.226 ± 0.065
3.382 ± 0.161
3.606 ± 0.196
2.578 ± 0.074
4.552 ± 0.265
3.433 ± 0.169
0.141 ± 0.003
1
B
30
1.029 ± 0.046
5.825 ± 0.314
4.007 ± 0.160
2.188 ± 0.034
5.825 ± 0.314
2.822 ± 0.070
0.952 ± 0.055
1
C
23
1.207 ± 0.040
1.731 ± 0.061
2.851 ± 0.061
1.947 ± 0.044
4.011 ± 0.200
2.932 ± 0.069
0.240 ± 0.010
1
D
47
1.000 ± 0.020
2.054 ± 0.040
2.402 ± 0.112
1.503 ± 0.039
3.614 ± 0.141
2.280 ± 0.090
0.197 ± 0.006
1
E
48
1.374 ± 0.025
2.767 ± 0.048
4.172 ± 0.118
2.423 ± 0.049
6.699 ± 0.191
3.761 ± 0.110
0.259 ± 0.010
1
F
28
1.092 ± 0.042
1.597 ± 0.400
2.060 ± 0.079
1.541 ± 0.048
3.439 ± 0.127
2.249 ± 0.078
0.194 ± 0.003
1
G
27
1.235 ± 0.036
2.205 ± 0.071
3.504 ± 0.427
2.013 ± 0.044
4.182 ± 0.259
2.625 ± 0.076
0.485 ± 0.027
1
H
18
1.142 ± 0.050
1.704 ± 0.061
2.825 ± 0.047
1.989 ± 0.040
4.240 ± 0.245
2.962 ± 0.084
0.244 ± 0.009
2.830 ± 0.271
I
31
1.287 ± 0.032
2.538 ± 0.081
3.424 ± 0.071
2.170 ± 0.031
5.935 ± 0.254
3.142 ± 0.096
0.365 ± 0.044
5.060 ± 0.596
J
17
1.381 ± 0.039
2.996 ± 0.081
3.520 ± 0.128
2.280 ± 0.065
7.174 ± 0.331
3.552 ± 0.129
0.105 ± 0.006
8.530 ± 0.944
K
19
1.207 ± 0.041
2.025 ± 0.075
3.018 ± 0.061
1.931 ± 0.047
4.098 ± 0.268
2.879 ± 0.096
0.377 ± 0.030
2.950 ± 0.209
L
42
0.969 ± 0.050
4.272 ± 0.241
3.014 ± 0.136
1.755 ± 0.056
4.272 ± 0.241
2.602 ± 0.071
0.026 ± 0.001
27.810 ± 1.985
n corresponds to the number of calls analysed within each category.
In the case of monosyllabic calls, the first two linear discriminant functions could account for 90.20% of the total variance (Fig. 1A, Table 2). The parameters F0min, F0max and Dur contributed mostly to LD1, while F0min, F5 and PF to LD2. As for multisyllabic calls, the first two linear discriminant functions accounted for 97.23% of the total variance (Fig. 1B, Table 3), with F0max and Fdom contributing mostly to LD1 while Fdom and F95 to LD2. The cross-validated DFA results showed that both the monosyllabic and multisyllabic calls were classified appropriately, with the correctness of 82.61% and 76.38% respectively (Appendix Tables S3 and S4).
Figure
1.
Discriminant Function Analysis (DFA) plot of the first two Linear Discriminants (LDs): (A) monosyllabic calls and (B) multisyllabic calls. Colour ellipse indicates 95% confidence interval. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
Table
2.
Linear discriminant functions (LD) of the discriminant function analysis (DFA) on monosyllabic calls and the proportion of total variance that each function could account for (Pr).
Table
3.
Linear discriminant functions (LD) of the discriminant function analysis (DFA) on multisyllabic calls and the proportion of total variance that each function could account for (Pr).
The PERMANOVA was performed to confirm the results, showing that the seven monosyllabic calls (PERMANOVA: F6,223 = 88.301, R2 = 0.704, P < 0.001) and five multisyllabic calls (PERMANOVA: F4,122 = 62.794, R2 = 0.673, P < 0.001) could be distinguished easily through the acoustic parameters. According to the pairwise comparisons, the differences between all pairs in monosyllabic calls were significant (P < 0.01). Similarly, all pairs within multisyllabic calls are significantly different (P < 0.01) except when comparing H and K (P = 0.628). We kept the categories the same as the prior definition (i.e., a total of twelve call categories), because of the clear differences in the spectrograms and contexts of H and K.
3.2
Context of call types
Context of 595 vocalisations was recorded and analysed. Generally, there was a strong correlation between call types and context (surroundings: χ2 = 447.534, df = 11, P < 0.001; behaviour of the signaller: χ2 = 1261.750, df = 55, P < 0.001; behaviour of the receiver: χ2 = 626.833, df = 558, P < 0.001). To be specific, B, F and I were emitted in the presence of intruders or predators, accompanied by the behaviour of alert and defence of nest and the response of defence of nest, approach or follow (Tables 4 and 5). A was involved with the behaviour of rest and copulation and the response of food provision and copulation (Tables 4 and 5); C and H with the behaviour of movement and individual alone and the response of approach and follow (Tables 4 and 5); D with a broad context of behaviour and response (Tables 4 and 5); E with beg for food and food provision specifically (Tables 4 and 5); G, K and J with the behaviour of rest and movement and the response of approach and follow (Tables 4 and 5); L with rest and play (Tables 4 and 5).
Table
4.
The context of the twelve call types of Azure-winged Magpies.
presence of intruders (49%); presence of predators (47%)
alert (26%); defence of nest (70%)
approach (46%); defence of nest (44%)
C
absence of danger (100%)
movement (53%); individual alone (26%)
approach (57%); follow (37%)
D
absence of danger (62%); presence of intruders (38%)
rest (20%); movement (21%); defence of nest (28%)
follow (18%); alert (23%); defence of nest (44%)
E
absence of danger (100%)
beg for food (88%)
approach (47%); food provision (53%)
F
presence of intruders (90%)
alter (21%); defence of nest (69%)
approach (34%); defence of nest (34%)
G
absence of danger (92%)
rest (27%); movement (69%)
approach (38%); follow (62%)
H
absence of danger (100%)
movement (34%); individual alone (66%)
approach (41%); follow (34%)
I
presence of intruders (57%)
movement (17%); alter (24%); defence of nest (43%)
follow (37%); alert (23%); defence of nest (20%)
J
absence of danger (97%)
rest (45%); movement (39%)
rest (24%); follow (59%)
K
absence of danger (82%)
rest (34%); movement (66%)
approach (42%); follow (52%)
L
absence of danger (100%)
rest (44%); play (56%)
rest (24%); play (28%); follow (28%)
The context included surroundings, behaviour of the signaller and behaviour of the receiver. % indicates the frequency with which a context is associated with a particular vocalisation. Only the top three contexts which accounted for more than 10% were listed.
This type is a quick, soft and high-pitched sound emitted by both juveniles and adults (Fig. 2A). The sound was usually produced after copulation, food provision or during resting and playing. Therefore, we suppose that this sound would be emitted when the basic requirements of the individual were met and represented a positive state.
Figure
2.
Spectrograms of call types of Azure-winged Magpie. Monosyllabic calls: A–G. Multisyllabic calls: H–L.
This is a noisy sound with great high F0max (5.825 ± 0.314 kHz) and long duration (0.952 ± 0.055 s) produced by individuals of all ages if any potential or real dangers happen (Fig. 2B). The responses varied, however, from an obvious response such as approaching the signaller and producing the same sound to the source of danger, to a slight response like neglecting the signaller. So, it may be a general alert and would recruit peers to mob only when enough hints pointed to the real danger.
3.3.3
C
This sound was emitted in flight and may play a role in sharing the location with the group members when individuals altered their location or sought contact with group members when the signaller was separated from the group (Fig. 2C). In response to this call, individuals would emit the same sound and some may approach the signaller. It seldom occurs alone and mainly consists of the multisyllabic call H as the first element.
3.3.4
D
This is a soft and low-frequency sound, with F0min only 1 kHz (1.000 ± 0.020 kHz; Fig. 2D). This category of sounds relates to a rather broad context and may be a basic contact call to keep in touch and enhance group cohesion.
3.3.5
E
E is a typical begging call with a wide bandwidth (4.276 ± 0.168 kHz; Fig. 2E). Nestlings and juveniles would produce the call with their beaks wide open when adults approached, accompanied by a flapping of wings. The incubating females would vocalise the sound as well to beg for food from their returned mates or helpers. The spectral view of this sound would dynamically change according to the urgency of the signaller.
3.3.6
F
This is a downslur sound in response to intrusion or a slight threat (Fig. 2F). When there was an intruder nearby, the individuals near the nest would continuously emit this sound. Only once was it recorded during the food competition.
3.3.7
G
This is a loud pre-flight call (Fig. 2G). Individuals would produce this sound before taking flights. However, the flight may not take off immediately after the vocalisation if there was no response from other group members.
3.3.8
H
As a multisyllabic call, it is composed of three elements on average (2.830 ± 0.271), with C consistently being the first element (Fig. 2H). Similar to C, this sound was produced when individuals were separated from the group. Adults would mainly emit this sound during flight (seeking group members) and approach the responder if exists, while fledglings would stay on the branch waiting for parents to find them.
3.3.9
I
I is another alert call in response to immediate danger and would initiate a chorus wherein most group members are involved after the first signaller (Fig. 2I). It was also recorded when no obvious danger was found by the observer and in this condition neither the signaller would be responded to nor a chorus would be initiated. Different from B, this alert seemed to target predators high in the air distinctively, such as a Eurasian sparrowhawk.
3.3.10
J
This sound is also a chorus call, consisting of eight elements on average (8.530 ± 0.944) (Fig. 2J). It was commonly given after a flight. Thus, we supposed that the sound may be involved with location recognition. When a valuable place is reached, whether for roosting or foraging, the chorus would be given by most members and the group would stay at the location for a period of time.
3.3.11
K
This multisyllabic call is composed of three elements on average (2.950 ± 0.209; Fig. 2K). Similar to the first element G, it was associated with flight. Once approached, followed and responded to by group members with the same vocalisation, a synchronised movement of the group would initiate.
3.3.12
L
This sound is a sequence of clicks with the interval less than 0.1 s (Fig. 2L). Each click lasts only 0.026 ± 0.001 s and it is hard to recognise the harmonics on the spectrogram. The sound was only produced by adults during resting. The sound would sometimes initiate the same vocalisation from other group members, but no direct connection was found.
4.
Discussion
In this study, twelve types of vocalisations were identified, seven of which were monosyllabic calls and the rest were multisyllabic calls consisting of a combination of different monosyllabic calls or a repetition of a single monosyllabic call. All categories were confirmed by DFA and PERMANOVA. A current study on 821 species of birds from 97 distinct families reported that birds have a mean repertoire of 7.25 distinct vocalisations, with the median value being 6 (Leighton and Birmingham, 2021). Therefore, the communication system of Azure-winged Magpies is more complex than people thought before, which could be an adaptation to their cooperative breeding strategy. As corroborated by Leighton (2017), the evolution of cooperative breeding was directly associated with increasing repertoire sizes. Azure-winged Magpies are prosocial and would help other individuals during both the breeding season and non-breeding season at high rates (Komeda et al., 1987; Horn et al., 2016). It is unsurprising that a large repertoire size is needed to maintain group cohesion along daily behaviour. Meanwhile, communication complexity was also shown to be proportional to the cognitive abilities of the species (Romani et al., 2022). The family Corvidae is comprised of species known for their innovative behaviour (Delehanty, 2022) and many of them have been reported to have a complex communication system. For example, Common Ravens (Corvus corax) have at least 18 distinct call types (Conner, 1985) and Blue Jays (Cyanocitta cristata) have 36 distinct vocalisations (Brewer, 2022). Azure-winged Magpies have also been shown to have a great talent in tool use and objective permanence (Wang et al., 2021; Zhang et al., 2021). Therefore, a complicated and nuanced vocal repertoire is indispensable for Azure-winged Magpies to efficiently transfer information and respond to conspecifics.
A strong connection was found between the vocalisation category and context. For instance, B and I were emitted when a potential predator was identified and E was associated with begging for food distinctively. However, the connection was not tight enough to induce a specific response of signal perceivers, which was predicted by 'functional referential signals' (Macedonia and Evans, 1993; Evans, 1997). Even if the attention is moved from the overall vocal repertoire to the alarm call, no direct evidence of functional referential signals could be found. B and I were vocalised to terrestrial and aerial predators respectively, similar to the textbook example of functional reference (Struhsaker, 1967). Nevertheless, they were given in other contexts as well and would elicit various responses. Similar phenomena were found in other species as well (Fichtel and Kappeler, 2002; Wheeler, 2010), with the terrestrial alarm calls not being context-specific whereas inducing terrestrial predator-specific responses in the playback experiments. It seemed that the functional reference could not give a comprehensive explanation of animal vocalisations. The narrow focus on functional referential signals may hold back the process of further investigation on other unspecific vocalisations (Wheeler and Fischer, 2012).
The limits of functional referential signals have been noticed gradually (Wheeler and Fischer, 2012). In order to discuss the referential signals from a broader aspect, a developed framework was given by Manser (2016). Two additional types of referents were recognised in the framework apart from the typical definition, external objects and events. One referent is individual traits and the social category. One call could carry information on individual recognition, individual status, social rank and social categories. For example, the threat-grunt-scream sequence in baboons could provide the dominance hierarchy of both the signallers and the opponents, with group members behaving surprisingly if the wrong sequence was given in the playback experiments (Bergman et al., 2003). The other referent is behavioural context. The current behaviour of the signaller could be reflected by specific call types, with E (begging call) being directly associated with food requirements. In other species, referential food calls would be emitted to attract mates, recruit group members and provide food for fledglings (Smith, 2017). Besides, the emotional state of the signaller could be included as well, as the sharp screams produced by Eurasian Griffon Vulture (Gyps fulvus) were always involved with stress (Manser, 2016). With more referents taken into consideration, researchers could discuss the underlying cognitive mechanisms from different references and investigate animal vocalisations from a broader perspective.
With respect to the alarm calls, three out of twelve (B, F and I) were found to be associated with vigilance. According to the previous study on 253 bird species, cooperative breeders would have a higher proportion of vocalisations related to vigilance (Leighton, 2017), which corresponds with what we found in Azure-winged Magpies. The results also showed that alarm calls of Azure-winged Magpies could be used in a broad context and induce various responses of receivers. This implies a complexity of information transferring within alarm calls. As mentioned before, I is an aerial predator-specific call. A typical aerial predator-specific alarm call would have a narrow bandwidth and high-pitched frequency so that the call would not travel too far to attract the attention of the predator (Marler, 2008). On the contrary, F is wideband and has a longer duration. This may result from their relatively large body size and aggressive property, which is similar to the research on Steller's Jays (Cyanocitta stelleri; Hope, 1980). Besides, B is a mobbing call given to a terrestrial predator. Many species have been reported to use different calls when encountering different types of predators (Smith, 2017). By being informed about the category of predators by totally different calls, receivers are allowed to choose an appropriate escape strategy quickly, thus increasing the survival rate (Marler, 1967). Meanwhile, the level of urgency is supposed to be encoded within these alarm calls. As found in Suricate (Suricata suricatta), calls would be noisier and lose their harmonic structure under high urgency (Manser, 2001). By recognising the nuanced differences in the alarm call, one could neglect the signaller and spare its time and energy when the situation is not emergent.
Several vocalisations related to flight were identified in Azure-winged Magpies as well. C and H were used to reunite left-behind members while G and K could induce the group to move from the original place to a new foraging or resting location. Similar findings were reported in Jungle Babbler (Argya striata), with the flight call inducing a synchronised movement (Yambem et al., 2021), and Pied Babbler (Turdoides bicolor), with 'purrs' urging group members to move (Engesser et al., 2017). Though reported in many birds, little attention was paid to its connection with the decision-making process of bird flocks. In Canada Goose (Branta canadensis; Raveling, 1969) and Mountain Gorilla (Gorilla gorilla berengei; Harcourt and Stewart, 1994), individuals have been observed to produce more moving calls to indicate their motivation to depart as well as assess the motivation of other group members. Therefore, it is a promising project to further investigate the moving calls of birds and discuss their potential as an essential process in the moving decision, along with the individual status of the first signaller.
5.
Conclusion
Generally, this study has given a basic but detailed description of the vocal repertoire of Azure-winged Magpies. A complex communication system is important for gregarious birds to coordinate group behaviours and respond to external events. The vocalisation category was context-associated and corresponded to the developed framework of 'functional referential signals'. The alarm calls of Azure-winged Magpies may include the category of predators and the urgency of taking action, but further studies are needed including playback experiments. This study could lay the foundation for all future investigations based on the vocal communication of this species.
Authors' contributions
ZL conceived and designed the experiments, XW and FY collected the data. XW and HS performed the analyses. XW wrote the first draft of the paper. FY, HS and ZL revised the manuscript. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We would like to thank Jieyun Li and Bin Qi for assistance with the preparation of this study. This work was supported by the National Key Research and Development Program of China (2022YFC3202104) and Natural Science Foundation of Jiangsu Province, China (BK20211151).
This work was conducted and approved by the Animal Welfare and Ethical Review Committee of Nanjing University (IACUC-2003001). All applicable international, national and institutional guidelines for the care and use of animals were followed. No animals were captured in this study.
Ambrosini, R., Saino, N., 2010. Environmental effects at two nested spatial scales on habitat choice and breeding performance of barn swallow. Evol. Ecol. 24, 491-508.
Ambrosini, R., Ferrari, R.P., Martinelli, R., Romano, M., Saino, N., 2006. Seasonal, meteorological, and microhabitat effects on breeding success and offspring phenotype in the barn swallow, Hirundo rustica. Ecoscience 13, 298-307.
Badgley, C., Fox, D.L., 2000. Ecological biogeography of North American mammals: species density and ecological structure in relation to environmental gradients. J. Biogeogr. 27, 1437-1467.
Barber, J.R., Crooks, K.R., Fristrup, K.M., 2010. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 25, 180-189.
Barton, K., 2022. MuMIn: multi-model inference. R package, version 1.46.0. .
Both, C., Artemyev, A.V., Blaauw, B., Cowie, R.J., Dekhuijzen, A.J., Eeva, T., et al., 2004. Large-scale geographical variation confirms that climate change causes birds to lay earlier. Proc R. Soc. B-Biol Sci. 271, 1657-1662.
Brans, K.I., Govaert, L., Engelen, J.M.T., Gianuca, A.T., Souffreau, C., De Meester, L., 2017. Eco-evolutionary dynamics in urbanized landscapes: evolution, species sorting and the change in zooplankton body size along urbanization gradients. Philos. Trans. R. Soc. B-Biol Sci. 372, 20160030.
Caizergues, A.E., Gregoire, A., Charmantier, A., 2018. Urban versus forest ecotypes are not explained by divergent reproductive selection. Proc. R. Soc. B-Biol Sci. 285, 20180261.
Casasole, G., Raap, T., Costantini, D., AbdElgawad, H., Asard, H., Pinxten, R., et al., 2017. Neither artificial light at night, anthropogenic noise nor distance from roads are associated with oxidative status of nestlings in an urban population of songbirds. Comp. Biochem. Physiol. Integr Physiol. 210, 14-21.
Chamberlain, D.E., Cannon, A.R., Toms, M.P., Leech, D.I., Hatchwell, B.J., Gaston, K.J., 2009. Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151, 1-18.
Ciach, M., Fröhlich, A., 2017. Habitat type, food resources, noise and light pollution explain the species composition, abundance and stability of a winter bird assemblage in an urban environment. Urban Ecosyst. 20, 547-559.
Clarke, K.R., 1993. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117-143.
Crain, C.M., Kroeker, K., Halpern, B.S., 2008. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 11, 1304-1315.
Crick, H.Q.P., Dudley, C., Glue, D.E., Thomson, D.L., 1997. UK birds are laying eggs earlier. Nature, 388, 526.
Da Silva, A., Samplonius, J.M., Schlicht, E., Valcu, M., Kempenaers, B., 2014. Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behav. Ecol. 25, 1037-1047.
de Jong, M., van den Eertwegh, L., Beskers, R.E., de Vries, P.P., Spoelstra, K., Visser, M.E., 2018. Timing of avian breeding in an urbanised world. Ardea 106, 31.
De Laet, J., Summers-Smith, J.D., 2007. The status of the urban House Sparrow Passer domesticus in north-western Europe: A review. J. Ornithol. 148, 275-278.
de Satgé, J., Strubbe, D., Elst, J., De Laet, J., Adriaensen, F., Matthysen, E., 2019. Urbanisation lowers great tit Parus major breeding success at multiple spatial scales. J. Avian Biol. 50, e02108.
Dominoni, D.M., Partecke, J., 2015. Does light pollution alter daylength? A test using light loggers on free-ranging European blackbirds (Turdus merula). Philos. Trans. R. Soc. B-Biol. Sci. 370, 20140118.
Dominoni, D., Quetting, M., Partecke, J., 2013. Artificial light at night advances avian reproductive physiology. Proc. R. Soc. B-Biol. Sci. 280, 20123017.
Dor, R., Safran, R.J., Sheldon, F.H., Winkler, D.W., Lovette, I.J., 2010. Phylogeny of the genus Hirundo and the Barn Swallow subspecies complex. Mol. Phylogenet. Evol. 56, 409-418.
Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al., 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27-46.
Fox, J., Weisberg, S., Adler, D., Bates, D., Baud-Bovy, G., Ellison, S., et al., 2012. Package 'car. '. R Foundation for Statistical Computing, Vienna.
García-Navas, V., Sanz, J.J., 2011. Seasonal decline in provisioning effort and nestling mass of Blue Tits Cyanistes caeruleus: experimental support for the parent quality hypothesis: Seasonal decline in parental care in Blue Tits. Ibis 153, 59-69.
Gaston, K.J., 2018. Lighting up the nighttime. Science 362, 744-746.
Grüebler, M.U., Naef-Daenzer, B., 2008. Fitness consequences of pre- and post-fledging timing decisions in a double-brooded passerine. Ecology 89, 2736-2745.
Grüebler, M.U., Korner-Nievergelt, F., Von Hirschheydt, J., 2010. The reproductive benefits of livestock farming in barn swallows Hirundo rustica: quality of nest site or foraging habitat?: Benefits of livestock farming. J. Appl. Ecol. 47, 1340-1347.
Guo, F., Bonebrake, T.C., Dingle, C., 2016. Low frequency dove coos vary across noise gradients in an urbanized environment. Behav. Process 129, 86-93.
Halfwerk, W., Holleman, L.J.M., Lessells, C.K.M., Slabbekoorn, H., 2011. Negative impact of traffic noise on avian reproductive success: Traffic noise and avian reproductive success. J. Appl. Ecol. 48, 210-219.
Håstad, O., Ödeen, A., 2014. A vision physiological estimation of ultraviolet window marking visibility to birds. PeerJ 2, e621.
Henseler, J., Ringle, C.M., Sinkovics, R.R., 2009. The use of partial least squares path modeling in international marketing. In: Sinkovics, R.R., Ghauri, P.N. (Eds. ), Advances in International Marketing. Emerald Group Publishing Limited, pp. 277-319. . (Accessed 10 Feb 2022).
Huchler, K., Schulze, C.H., Gamauf, A., Sumasgutner, P., 2020. Shifting breeding phenology in Eurasian Kestrels Falco tinnunculus: Effects of weather and urbanization. Front. Ecol. Evol. 8, 247. .
Huet des Aunay, G., Grenna, M., Slabbekoorn, H., Nicolas, P., Nagle, L., Leboucher, G., et al., 2017. Negative impact of urban noise on sexual receptivity and clutch size in female domestic canaries. Ethology 123, 843-853.
Imhoff, M.L., Zhang, P., Wolfe, R.E., Bounoua, L., 2010. Remote sensing of the urban heat island effect across biomes in the continental USA. Remote Sens. Environ. 114, 504-513.
Injaian, A.S., Poon, L.Y., Patricelli, G.L., 2018. Effects of experimental anthropogenic noise on avian settlement patterns and reproductive success. Behav. Ecol. 29, 1181-1189.
Jokimäki, J., Suhonen, J., Vuorisalo, T., Kövér, L., Kaisanlahti-Jokimäki, M. -L., 2017. Urbanization and nest-site selection of the Black-billed Magpie (Pica pica) populations in two Finnish cities: From a persecuted species to an urban exploiter. Lands Urban Plan. 157, 577-585.
Jones, T., Cresswell, W., 2010. The phenology mismatch hypothesis: are declines of migrant birds linked to uneven global climate change? J. Anim. Ecol. 79, 98-108.
Kight, C.R., Swaddle, J.P., 2011. How and why environmental noise impacts animals: an integrative, mechanistic review: Environmental noise and animals. Ecol. Lett. 14, 1052-1061.
Kight, C.R., Saha, M.S., Swaddle, J.P., 2012. Anthropogenic noise is associated with reductions in the productivity of breeding Eastern Bluebirds (Sialia sialis). Ecol. Appl. 22, 1989-1996.
Kosiński, Z., 2001. Effects of urbanization on nest site selection and nesting success of the Greenfinch Carduelis chloris in Krotoszyn, Poland. Ornis Fenn. 78, 175-183.
Lê, S., Josse, J., Husson, F., 2008. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 25, 1-18.
McKinney, M.L., 2008. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 11, 161-176.
Merckx, T., Souffreau, C., Kaiser, A., Baardsen, L.F., Backeljau, T., Bonte, D., et al., 2018. Body-size shifts in aquatic and terrestrial urban communities. Nature 558, 113-116.
Moeller, A.P., 2010. The fitness benefit of association with humans: elevated success of birds breeding indoors. Behav, Ecol. 21, 913-918.
Møller, A.P., Rubolini, D., Lehikoinen, E., 2008. Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc. Natl. Acad. Sci. 105, 16195-16200.
Møller, A.P., Díaz, M., Grim, T., Dvorská, A., Flensted-Jensen, E., Ibáñez-Álamo, J., et al., 2015. Effects of urbanization on bird phenology: A continental study of paired urban and rural populations. Clim. Res. 66, 185-199.
Nebel, S., Mills, A., McCracken, J.D., Taylor, P.D., 2010. Declines of aerial insectivores in North America follow a geographic gradient. Avian Conserv. Ecol. 5, 1.
Oksanen, J., Kindt, R., Legendre, P., O'Hara, B., Stevens, M.H.H., Oksanen, M.J., et al., 2007. The vegan package. Commun. Ecol. Pack. 10, 719.
Ouyang, J.Q., de Jong, M., van Grunsven, R.H.A., Matson, K.D., Haussmann, M.F., Meerlo, P., et al., 2017. Restless roosts: Light pollution affects behavior, sleep, and physiology in a free-living songbird. Glob. Change Biol. 23, 4987-4994.
Pagani-Núñez, E., Uribe, F., Hernández-Gómez, S., Muñoz, G., Senar, J.C., 2014. Habitat structure and prey composition generate contrasting effects on carotenoid-based coloration of great tit Parus major nestlings: determinants of nestling coloration. Biol. J. Linn. Soc. 113, 547-555.
Pagani-Nunez, E., He, C., Li, B., Li, M., He, R., Jiang, A., et al., 2016. The breeding ecology of the barn swallow Hirundo rustica gutturalis in South China. J. Trop. Ecol. 32, 260-263.
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., Team, R.C., 2007. Linear and nonlinear mixed effects models. R Package Version 3, 1-89.
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. .
Raja-aho, S., Eeva, T., Suorsa, P., Valkama, J., Lehikoinen, E., 2017. Juvenile Barn Swallows Hirundo rustica L. from late broods start autumn migration younger, fuel less effectively and show lower return rates than juveniles from early broods. Ibis 159, 892-901.
Ramirez, F., Coll, M., Navarro, J., Bustamante, J., Green, A.J., 2018. Spatial congruence between multiple stressors in the Mediterranean Sea may reduce its resilience to climate impacts. Sci. Rep. 8, 14871.
Schlesinger, M.D., Manley, P.N., Holyoak, M., 2008. Distinguishing stressors acting on land bird communities in an urbanizing environment. Ecology 89, 2302-2314.
Senzaki, M., Barber, J.R., Phillips, J.N., Carter, N.H., Cooper, C.B., Ditmer, M.A., et al., 2020. Sensory pollutants alter bird phenology and fitness across a continent. Nature 587, 605-609.
Seress, G., Hammer, T., Bókony, V., Vincze, E., Preiszner, B., Pipoly, I., et al., 2018. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird. Ecol. Appl. 28, 1143-1156.
Seress, G., Sándor, K., Evans, K.L., Liker, A., 2020. Food availability limits avian reproduction in the city: An experimental study on great tits Parus major. J. Anim. Ecol. 89, 1570-1580.
Seto, K.C., Fragkias, M., Güneralp, B., Reilly, M.K., 2011. A meta-analysis of global urban land expansion. PLoS One 6, e23777.
Shannon, G., McKenna, M.F., Angeloni, L.M., Crooks, K.R., Fristrup, K.M., Brown, E., et al., 2016. A synthesis of two decades of research documenting the effects of noise on wildlife: Effects of anthropogenic noise on wildlife. Biol. Rev. 91, 982-1005.
Sievers, M., Hale, R., Parris, K.M., Swearer, S.E., 2018. Impacts of human-induced environmental change in wetlands on aquatic animals: Animal communities, populations and individuals in human-impacted wetlands. Biol. Rev. 93, 529-554.
Sol, D., Lapiedra, O., González-Lagos, C., 2013. Behavioural adjustments for a life in the city. Anim. Behav. 85, 1101-1112.
Sprau, P., Mouchet, A., Dingemanse, N.J., 2017. Multidimensional environmental predictors of variation in avian forest and city life histories. Behav. Ecol. 28, 59-68.
Stoleson, S.H., Beissinger, S.R., 1999. Egg viability as a constraint on hatching synchrony at high ambient temperatures. J. Anim. Ecol. 68, 951-962.
Strasser, E.H., Heath, J.A., 2013. Reproductive failure of a human-tolerant species, the American kestrel, is associated with stress and human disturbance. J. Appl. Ecol. 50, 912-919.
Summers-Smith, J.D., 2003. The decline of the House Sparrow: A review. Br. Birds 96, 439-446.
Symonds, M.R.E., Moussalli, A., 2011. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike's information criterion. Behav. Ecol. Sociobiol. 65, 13-21.
Teglhøj, P.G., 2017. A comparative study of insect abundance and reproductive success of barn swallows Hirundo rustica in two urban habitats. J. Avian Biol. 48, 846-853.
Tuanmu, M. -N., Jetz, W., 2014. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling: Consensus land cover. Glob. Ecol. Biogeogr. 23, 1031-1045.
Verhulst, S., Nilsson, J. -Å., 2008. The timing of birds' breeding seasons: a review of experiments that manipulated timing of breeding. Philos. Trans. R. Soc. B-Biol. Sci. 363, 399-410.
Visser, M.E., Holleman, L.J.M., Gienapp, P., 2006. Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia 147, 164-172.
Wang, J. -S., Tuanmu, M. -N., Hung, C. -M., 2021. Effects of artificial light at night on the nest-site selection, reproductive success and behavior of a synanthropic bird. Environ. Pollut. 288, 117805.
Welbers, A.A.M.H., van Dis, N.E., Kolvoort, A.M., Ouyang, J., Visser, M.E., Spoelstra, K., et al., 2017. Artificial light at night reduces daily energy expenditure in breeding Great Tits (Parus major). Front. Ecol. Evol. 5, 55.
World Health Organization (Ed. ), 1999. The World Health Report 1999: making a difference. WHO, Geneva.
Xu, Y., Cao, Z., Wang, B., 2020. Effect of urbanization intensity on nest-site selection by Eurasian Magpies (Pica pica). Urban Ecosyst. 23, 1099-1105.
Zhang, K., Wang, R., Shen, C., Da, L., 2010. Temporal and spatial characteristics of the urban heat island during rapid urbanization in Shanghai, China. Environ. Monit. Assess. 169, 101-112.
Zhao, Y., Liu, Y., Scordato, E.S.C., Lee, M., Xing, X., Pan, X., et al., 2021. The impact of urbanization on body size of Barn Swallows Hirundo rustica gutturalis. Ecol. Evol. 11, 612-625.
Zink, R.M., Pavlova, A., Rohwer, S., Drovetski, S.V., 2006. Barn swallows before barns: Population histories and intercontinental colonization. Proc. R. Soc. B-Biol. Sci. 273, 1245-1251.
Zuur, A.F., Ieno, E.N., Walker, N., Saveliev, A.A., Smith, G.M., 2009. Mixed effects models and extensions in ecology with R. Springer, New York. . (Accessed 1 Feb 2022).
Table
2.
Linear discriminant functions (LD) of the discriminant function analysis (DFA) on monosyllabic calls and the proportion of total variance that each function could account for (Pr).
Table
3.
Linear discriminant functions (LD) of the discriminant function analysis (DFA) on multisyllabic calls and the proportion of total variance that each function could account for (Pr).
presence of intruders (49%); presence of predators (47%)
alert (26%); defence of nest (70%)
approach (46%); defence of nest (44%)
C
absence of danger (100%)
movement (53%); individual alone (26%)
approach (57%); follow (37%)
D
absence of danger (62%); presence of intruders (38%)
rest (20%); movement (21%); defence of nest (28%)
follow (18%); alert (23%); defence of nest (44%)
E
absence of danger (100%)
beg for food (88%)
approach (47%); food provision (53%)
F
presence of intruders (90%)
alter (21%); defence of nest (69%)
approach (34%); defence of nest (34%)
G
absence of danger (92%)
rest (27%); movement (69%)
approach (38%); follow (62%)
H
absence of danger (100%)
movement (34%); individual alone (66%)
approach (41%); follow (34%)
I
presence of intruders (57%)
movement (17%); alter (24%); defence of nest (43%)
follow (37%); alert (23%); defence of nest (20%)
J
absence of danger (97%)
rest (45%); movement (39%)
rest (24%); follow (59%)
K
absence of danger (82%)
rest (34%); movement (66%)
approach (42%); follow (52%)
L
absence of danger (100%)
rest (44%); play (56%)
rest (24%); play (28%); follow (28%)
The context included surroundings, behaviour of the signaller and behaviour of the receiver. % indicates the frequency with which a context is associated with a particular vocalisation. Only the top three contexts which accounted for more than 10% were listed.

 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds