Johann H. Van Niekerk, Rodrigo Megía-Palma, Giovanni Forcina. 2022: Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa. Avian Research, 13(1): 100047. DOI: 10.1016/j.avrs.2022.100047
Citation:
Johann H. Van Niekerk, Rodrigo Megía-Palma, Giovanni Forcina. 2022: Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa. Avian Research, 13(1): 100047. DOI: 10.1016/j.avrs.2022.100047
Johann H. Van Niekerk, Rodrigo Megía-Palma, Giovanni Forcina. 2022: Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa. Avian Research, 13(1): 100047. DOI: 10.1016/j.avrs.2022.100047
Citation:
Johann H. Van Niekerk, Rodrigo Megía-Palma, Giovanni Forcina. 2022: Thermoregulatory function and sexual dimorphism of the throat sack in Helmeted Guineafowl (Numida meleagris) across Africa. Avian Research, 13(1): 100047. DOI: 10.1016/j.avrs.2022.100047
Department of Environmental Sciences, College of Agriculture and Environmental Sciences, University of South Africa, P.O. Box 392, Pretoria, 0003, South Africa
b.
Universidad de Alcalá (UAH), Department of Biomedicine and Biotechnology, School of Pharmacy, A-2 Highway, km 33.600, 28805, Alcalá de Henares, Madrid, Spain
c.
Universidad de Alcalá (UAH), Global Change Ecology and Evolution Research Group (GloCEE), Departamento de Ciencias de la Vida, 28805, Alcalá de Henares, Madrid, Spain
d.
CIBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, InBIO Laboratório Associado, Campus de Vairão, Universidade do Porto, 4485-661, Vairão, Portugal
e.
BIOPOLIS Program in Genomics, Biodiversity and Land Planning, CIBIO, Campus de Vairão, 4485-661, Vairão, Portugal
Funds:
the Portuguese Foundation for Science and TechnologyPTDC/BAA-AGR/28866/2017
the Portuguese Foundation for Science and TechnologyCEECIND/04084/2017
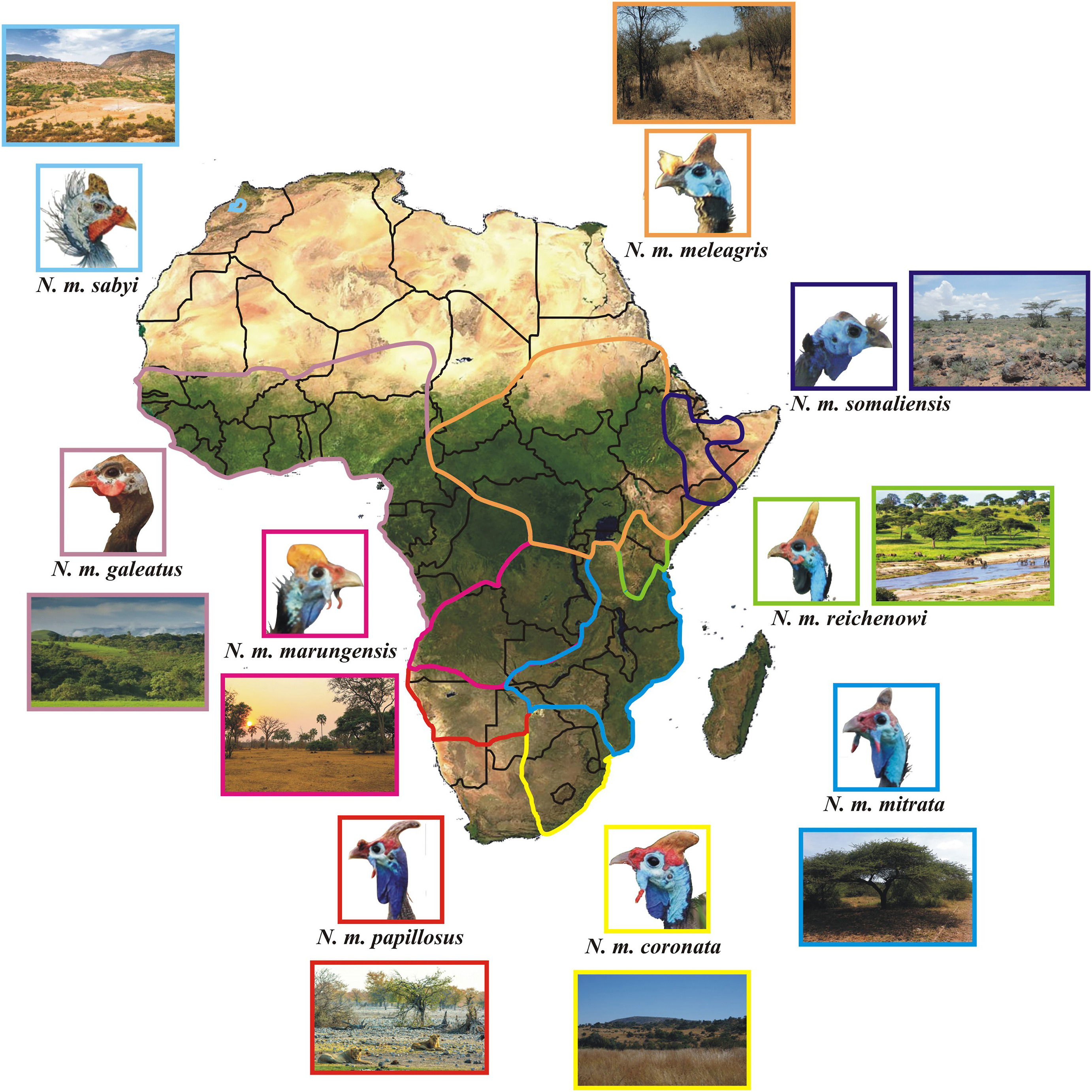
The responses of ground-dwelling birds to heat and cold stress encompass a variety of behavioural, physiological and even morphological mechanisms. However, the role of glabrous skin in this respect has been marginally addressed so far. The Helmeted Guineafowl (Numida meleagris) is a landfowl distributed across Sub-Saharan Africa with eight traditionally recognised extant subspecies. Among the most prominent morphological traits underlying intraspecific variability are size and pigmentation of the bare throat skin (or sack), which might be related to the different habitats and environmental conditions across its wide range. In order to explore the Helmeted Guineafowl range-wide sack variation and pigmentation in relation to thermoregulation and sexual signalling, we collected morphometric and environmental information for N. m. coronata integrating field data with the inspection of photographic material encompassing seven subspecies and environmental information from their habitats. Field data evidenced that sack size was significantly correlated with ambient temperature, thus pointing to a likely involvement of the throat sack in thermoregulation. When the pictorial data from all subspecies were pooled, sack size correlated negatively with biomass, rainfall and humidity, while a positive correlation was found with annual solar irradiation. Sack size correlated positively with monthly temperature variation among the blue-throated subspecies from southern Africa as opposed to the black-throated subspecies ranging north to Zambia and Mozambique. Still, in this latter group the sack was often larger during winter months, possibly to maximise solar radiation absorbance. Noteworthy, sack size was related to sex dimorphism in two subspecies. Sack morphology and colour in the Helmeted Guineafowl likely modulate body temperature by evaporative cooling or heating upon needs, but in some subspecies it is also seemingly related to sexual signalling. Additional studies are needed to fully understand the multifunctionality of this important morphological feature in this species.
The vast majority of known birds, extant or fossil, fall into the clades Ornithuromorpha and Enantiornithes, which together form the Ornithothoraces. Ornithuromorpha includes all extant birds and their close relatives, while Enantiornithes is an exclusively Cretaceous clade of morphologically primitive taxa. In this paper, we describe as "basal birds" the relatively small number of even more primitive Mesozoic avians that fall outside Ornithothoraces. Perennial interest in the origin of birds and the early evolution of flight has made basal birds a focus of research activity in paleontology ever since 1861 when the first basal bird, Archaeopteryx lithographica, was discovered. After about a century and a half of research, the known roster of basal birds includes nine described genera: Archaeopteryx, Jeholornis, Zhongornis, Sapeornis, Zhongjianornis, Eoconfuciusornis, Confuciusornis, Changchengornis and Jinzhouornis. All of these genera other than Archaeopteryx are from China, although a probable confuciusornithid is also known to be from North Korea (Paek and Kim, 1996; Gao et al., 2009). The phylogenetic relationships of basal birds have been studied for many years (Fig. 1).
Figure
1.
Phylogenetic relationships of basal birds (synthesized from Chiappe et al., 1999; Chiappe, 2002; Hou et al., 2002; Zhou and Zhang, 2002b; Zhang et al., 2008; Gao et al., 2008; Zhou et al., 2009)
For consistency with the paleontological literature, we number the digits of the basal bird manus as Ⅰ–Ⅲ in this paper, rather than Ⅱ–Ⅳ as a scheme of developmental homologies would suggest.
Stratigraphic and geochronological background
All of the Chinese basal birds that have so far been discovered come from two northern provinces, Liaoning and Hebei (Zhang et al., 2009). The basal bird-bearing deposits comprise the Yixian Formation and the overlying Jiufotang Formation in Liaoning, as well as the Dabeigou Formation in Hebei (Chang et al., 2003; Zhou and Zhang, 2006; Zhou, 2006; Zhang et al., 2008). The Dabeigou Formation is thought to be stratigraphically lower than the Yixian Formation (Zhou and Zhang, 2007). The age of the Jiufotang Formation is about 120 Mya (He et al., 2004), whereas the Yixian Formation is about 125 Mya old (Swisher et al., 2002) and the Dabeigou Formation about 131 Mya old (He et al., 2006).
The Chaomidianzi Formation was named in 1999 (Chiappe et al., 1999) in reference to a body of strata that lies below the Yixian Formation. A section at the village of Sihetun, south of Beipiao City (Fig. 2), was selected as the Chaomidianzi type section. However, the strata of the putative Chaomidianzi Formation were subsequently recognized as belonging to the Jianshangou Beds, which are basal deposits of the Yixian Formation (cf. Chen et al., 2005).
Figure
2.
Localities of various basal birds in China. Note that cities are focal points for large administrative areas that may contain several fossil localities.
Eight genera and twelve valid species of Chinese basal birds have been described in the last 15 years (Table 1), dramatically increasing our knowledge of early birds.
Table
1.
List of Chinese basal birds and their geographic and stratigraphic distribution. Holotype specimens are housed at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), Beijing; the Dalian Natural History Museum (D), Dalian, Liaoning Province; and the National Geological Museum of China (GMV), Beijing.
J. prima, the only known species of this genus, was briefly described by Zhou and Zhang (2002b) and later compared in detail to Archaeopteryx(Zhou and Zhang, 2003a; Fig. 3). The generic name is derived from the Jehol Biota whereas the specific name refers to the primitive nature of Jeholornis, as demonstrated by the long bony tail. Between them, the two publications described three specimens of J. prima.
Figure
3.
Holotype of Jeholornis prima, IVPP V13274
J. prima has been repeatedly found in phylogenetic analyses (Zhou and Zhang, 2002b; Chiappe and Dyke, 2006; Gao et al., 2008) to be the most primitive bird from China and can be easily distinguished from other Chinese fossil birds by the long skeletal tail consisting of more than 20 caudal vertebrae. J. prima also displays the following characters: large body size; teeth present; lacrimal "T" shaped; mandibles robust; first phalanx of third manual digit twice as long as second phalanx; lateral sternal trabecula penetrated by caudally located round fenestrae and not fused with body of sternum; length ratio of forelimb (humerus plus ulna plus carpometacarpus) to hind limb (femur plus tibiotarsus plus tarsometatarsus) about 1.2. Feather impressions are well preserved in the two referred specimens, although not in the holotype. One of the referred specimens preserves long, asymmetric wing feathers, while the other preserves tail feathers that have been interpreted as forming a more or less fan-shaped surface at the caudal end of the tail (Zhou and Zhang, 2002b).
Shenzhouraptor sinensis(Ji et al., 2002a) and Jixiangornis orientalis(Ji et al., 2002b) are both regarded as junior synonyms of J. prima on the basis of similar diagnostic features (Zhou and Zhang, 2006). For instance, S. sinensis and J. orientalis each have a long skeletal tail consisting of more than 20 caudal vertebrae, a character state also present in J. prima; the length ratio of the forelimb to hind limb is 1.27 in S. sinensis and 1.31 in J. orientalis, both comparable to the value of 1.2 seen in J. prima; in S. sinensis and J. orientalis the deltopectoral crest of the humerus is well developed, as in J. prima. The specimens assigned to S. sinensis and J. orientalis differ from J. prima in completely lacking teeth, but this difference may be an artifact of ontogeny and/or preservation given that only a few tiny teeth are present in J. prima itself.
The ecology of J. prima was also discussed in the papers cited above (Zhou and Zhang, 2002b, 2003a). Over 50 ovules are well preserved in the abdominal region of the holotype and the robust, nearly edentulous mandible and well-developed hyoid lend support to the inference that J. prima was a seed-eater. The ovules, when discovered, represented the first direct evidence of diet in a Mesozoic bird from China. J. prima displays some derived features, such as a strut-like coracoid, which imply a capacity for powered flight. J. prima is likely to have been arboreal, given its reversed hallux, long and curved pedal claws.
J. prima is an important taxon partly because the long skeletal tail, which resembles that of many dromaeosaurids in having some elongated prezygapophyses and chevrons and may suggest a relatively close phylogenetic relationship between birds and dromaeosaurids (Zhou and Zhang, 2002b).
Sapeornis
Sapeornis includes two species: S. chaoyangensis(Zhou and Zhang, 2002a, 2003b), which is the largest currently known basal bird, and S. angustis (Provini et al., 2009; Fig. 4). The genus was named for the Society of Avian Paleontology and Evolution (SAPE), which held its successful fifth conference in China in 2000. Characteristic features shared by both species of Sapeornis include: robust furcula with slim hypocleidum; forelimb extraordinarily elongated, about 1.5 times the length of the hind limb; proximal end of humerus with oval foramen and deltoid crest measuring about one-third of humeral length; third manual digit consisting of two slender phalanges in addition to the metacarpal.
Figure
4.
Holotype of Saperornis chaoyangensis, IVPP V12698
Didactylornis jii was described as a new sapeornithid bird and putatively distinguished from S. chaoyangensis by its differing manual and pedal phalangeal counts and proportions (Yuan, 2008). However, these apparent differences may easily result from poor preservation or damage and we consider D. jii to be a probable junior synonym of S. chaoyangensis, given that the holotype of D. jii appears to be a normal S. chaoyangensis specimen in almost all respects.
Sapeornis has been the subject of phylogenetic and ecological interpretations. One phylogenetic result (Zhou et al., 2008) shows that Sapeornis is the most basal bird genus with a fused pygostyle, a character indicating that Sapeornis is closer to extant birds than either A. lithographica or J. prima (but see Zhou et al., 2009). However, other analyses have indicated that Sapeornis may be more derived than Confuciusornis, which also presents a fused pygostyle (Gao et al., 2008; O'Connor et al., 2009). Under this interpretation, Sapeornis would represent the most derived known basal bird. It is not surprising that the relatively primitive Sapeornis shares some significant characters with Archaeopteryx and dromaeosaurids, including a short and non-strut-like coracoid and retention of the fifth metatarsal. The proportionally elongated forelimbs may indicate that the wings were long and suitable for soaring (Zhou and Zhang, 2002a, 2003b). Such an unusual aerodynamic capability would demonstrate considerable functional and ecological diversity among Early Cretaceous birds.
A few other features of the skeleton, particularly the pygostyle and hypocleidum, also support the inference that the two Sapeornis species were more anatomically specialized for flight than either Jeholornis or Archaeopteryx(Zhou and Zhang, 2003b). The opposable hallux, long pedal digit Ⅰ and highly curved unguals, suggest well-developed perching ability. The presence of gastroliths in S. chaoyangensis(Zhou and Zhang, 2003b) and the reduced teeth, imply that Sapeornis may have been herbivorous. Members of this genus can perhaps be pictured as large, arboreal, aerodynamically capable plant-eaters.
S. chaoyangensis
The specific name of S. chaoyangensis is derived from Chaoyang City (Fig. 2), the locality where the holotype specimen was collected. S. chaoyangensis lacks a suite of distinctive features seen in S. angustis(see below) and also seems to be substantially larger than S. angustis. However, the extent of the actual size discrepancy is uncertain, because S. angustis is based on a subadult individual.
S. angustis
S. angustis is a recently described species of Sapeornis (Provini et al., 2009); the specific name comes from the Latin word for narrow. The only known specimen is thought to be a subadult individual, based on the lack of preserved uncinate processes, the lack of a sternum and the fact that the sacral vertebrae are not fully fused into a synsacrum. Apart from a possibly smaller body size, S. angustis evidently differs from S. chaoyangensis in the following characters: maximum of 6 sacrals (compared to 7 in S. chaoyangensis); distinctively narrow humeral deltopectoral crest whose dorsodistal end does not form an acute prominence; furcular rami slightly narrower in proportion to their length; shorter hypocleidum; shorter pubic symphysis and relatively longer metacarpal Ⅰ (approximately one-third of the length of metacarpal Ⅱ, compared to approximately one-quarter in S. chaoyangensis).
Zhongornis
The generic name Zhongornis(Gao et al., 2008) means "intermediate bird" and the name of the only known species Z. haoae honors a Ms. Hao, who donated the only known specimen for scientific research. The specimen is distinguished by the following combination of features: teeth absent; tail consists of 13–14 unfused caudal vertebrae and no pygostyle is developed; coracoid strut-like and expanded proximally; deltopectoral crest of humerus lacks foramen; manual phalangeal formula 2-3-3-x-x; largest manual claw on digit Ⅰ, with unguals Ⅱ and Ⅲ subequal.
The 13–14 unfused caudal vertebrae of Zhongornis form a tail that is intermediate in structure between those of Jeholornis on the one hand and Sapeornis and Confuciusornis on the other. Jeholornis has a long tail consisting of more than 20 unfused caudal vertebrae, while Sapeornis and Confuciusornis have tails with fewer than ten unfused caudal vertebrae followed by a pygostyle. Gao et al. (2008) recovered Zhongornis as the sister taxon of a clade including Sapeornis, Confuciusornithidae and Ornithothoraces, but the results of their analysis were atypical in placing confuciusornithids basal to Sapeornis within this group. In the present paper the positions of Sapeornis and Zhongornis relative to more derived birds are treated as uncertain (Fig. 1).
Zhongjianornis
The generic and specific names of Z. yangi(Zhou et al., 2009) both honor the founder of Chinese vertebrate paleontology and of the Institute of Vertebrate Paleontology and Paleoanthropology (ⅣPP), the late Professor Zhongjian Yang (Chung-Chien Young). Zhongjianornis is a pigeon-sized bird with toothless jaws; its snout is pointed; its humerus has a large and robust deltopectoral crest that is as wide as the humeral shaft and more than one-third the total length of the humerus; its manual unguals are small and slightly curved; and in its pes metatarsal Ⅳ is longer than Ⅱ and Ⅲ (Fig. 5).
Zhou et al. (2009) also considered the ecology and phylogenetic position of Zhongjianornis. The presence of uncinate processes in the only fully-developed specimen may indicate that Zhongjianornis is more derived than Archaeopteryx, Jeholornis and Sapeornis, although the apparent lack of uncinate processes in these three taxa has been interpreted as resulting simply from incomplete ossification in described specimens (Provini et al., 2009). One of the most significant features of Zhongjianornis is the toothless jaws. It can be assumed that minimizing body mass is important in a flying bird and that reducing mass at a point far from the center of gravity could have a disproportionate benefit because of the potential gain in stability. Loss of the dentition could have been a response to this selection pressure. Similarly, the enlarged humeral deltopectoral crest may represent an adaptation for increased flight ability. Zhongjianornis is also likely to have been arboreal, judging by the long, curved pedal unguals and the proportions of the hind limb, including the pes.
Eoconfuciusornis
E. zhengi is the stratigraphically oldest basal bird so far discovered in China (Zhang et al., 2008). The generic name uses the Greek prefix "eo-", which means dawn, to modify the name of the related genus Confuciusornis. The specific name honors the famous Chinese ornithologist Guangmei Zheng. Eoconfuciusornis can easily be identified by the following features: lateral depressions in thoracic vertebrae weakly developed; scapula lacks prominent acromion and glenoid facet; coracoid short, with relatively wide sternal facet; deltopectoral crest at proximal end of humerus not prominent; proximal end of humerus less than twice as wide as distal end and lacks a foramen; astragalus perforated by several foramina; tarsometatarsus slightly more than half as long as tibia (Fig. 6).
Figure
6.
Holotype of Eoconfuciusornis zhengi, IVPP V11977
Eoconfuciusornis is more primitive than other known confuciusornithids (Zhang et al., 2008), a clade that also includes Confuciusornis, Changchengornis and Jinzhouornis. Eoconfuciusornis shares a number of important features with other confuciusornithids, including toothless jaws and a reduced ungual on manual digit Ⅱ. Given the basal phylogenetic position of Eoconfuciusornis within confuciusornithids, as well as its relatively great age, it is unsurprising that this taxon retains some primitive features, such as the unitary but poorly ossified and acarinate sternum. This condition is transitional between more primitive basal birds such as Jeholornis, which possesses a pair of flat sternal plates and the more derived confuciusornithid Confuciusornis which has a sternum with a carina (Zhang et al., 2008). This trend in sternal morphology may indicate a gradual increase in flight capability. Eoconfuciusornis is a highly significant taxon in that it records an intermediate stage in the continuum of sternal morphology and exemplifies the basal confuciusornithid anatomical condition in general.
Confuciusornis
Confuciusornis is a derived member of Confuciusornithidae. The genus is named after Confucius, the greatest thinker and educator in Chinese history and is distinguished by the following combination of features (Chiappe et al., 1999; Hou et al., 1999; Li et al., 2008; Zhang et al., 2008): teeth absent and horny bill present, as in other confuciusornithids; antorbital fenestra small and triangular; deltopectoral crest enlarged; manual ungual Ⅱ much smaller than manual unguals Ⅰ and Ⅲ; hallux not exceeding 50% of length of pedal digit Ⅱ. Confuciusornis is also the most diverse basal bird genus, containing three valid species: C. sanctus, C. dui and C. feducciai. Two additional nominal species, C. chuanzhous and C. suniae(Hou, 1997), are thought to be junior synonyms of C. sanctus(Chiappe et al., 1999). C. sanctus is the most abundant species, of which more than one thousand specimens have been collected (Zhang et al., 2009). Confuciusornis has a relatively long femur, short tarsometatarsus, well-developed fibula and reversed hallux, all of which are features suggestive of arboreality and at least in C. feducciai, the manual phalangeal proportions are such that the hand appears well adapted for grasping (Zhang et al., 2009). However, some features of C. sanctus have also been regarded as indicating partly terrestrial habits (Chiappe et al., 1999).
C. chuanzhous(Hou, 1997) was established based on a single specimen consisting only of a partial hind limb. The specimen was reported (Hou, 1997) to display the following features: tibia robust, with unexpanded distal end and associated feather impressions; calcaneus and astragalus separate; fifth metatarsal distinct from other metatarsals except at proximal end; pedal digit Ⅰ extremely small and less curved than in other early birds. However, Chiappe et al. (1999) argued convincingly that all of these supposedly diagnostic characters either were based on misinterpretations of the specimen or did not represent significant differences from C. sanctus, making C. chuanzhous synonymous with this taxon.
The one specimen assigned to C. suniae(Hou, 1997) has the following features: notch at rostral end of snout; parietal large; cervical vertebrae wide, with medium-sized lateral depressions and low, caudocranially short neural spines; thoracic vertebrae narrow and long, with elongate, deep lateral depressions; transverse processes of last three lumbar vertebrae fused; caudal vertebrae nearly fused. Although the putative C. suniae specimen is nearly complete, its morphology falls within the range of variation seen in C. sanctus, so C. suniae cannot be considered a valid species (Chiappe et al., 1999).
C. sanctus
C. sanctus was the first basal bird found in China and only the second basal bird found worldwide (Fig. 7). This species is now the best known in the genus Confuciusornis and is one of the most noteworthy fossil birds due to its transitional phylogenetic position (shared, of course, with other confuciusornithids) and to the large number of recovered specimens. Most C. sanctus specimens have come from the Yixian Formation. However, one published specimen has come from the Jiufotang Formation, indicating that this species persisted for at least 5 Mya (Dalsätt et al., 2006). C. sanctus displays the following features: dentary forms anteroventral expansion; oval foramen pierces deltopectoral crest of humerus; intermediate phalanx of manual digit Ⅱ strongly bowed (Hou et al., 1995a, 1995b, 1999; Martin et al., 1998; Chiappe et al., 1999).
Figure
7.
Forelimb part of the holotype of Confuciusornis sancus, IVPP V10918
C. sanctus has played an important role in the study of avian evolution, owing to its status as the first basal bird to be discovered after Archaeopteryx. As the first known confuciusornithid, C. sanctus bridged the morphological gap between Archaeopteryx and Ornithothoraces, shedding further light on the early radiation of birds. Following the discovery of Jeholornis and Sapeornis, Confuciusornis was no longer the most basal Chinese fossil bird genus. However, C. sanctus continues to play a crucial role in studies of Early Cretaceous birds, partly because such a large number of specimens are available.
C. sanctus has been interpreted as either a tree climber (Hou et al., 1995b) or a bird that was partly arboreal and partly cursorial (Chiappe et al., 1999). This species was also the first basal bird with toothless jaws and a horny bill to be discovered. A paper describing fish remains preserved in the neck region of a C. sanctus was published in 2006 (Dalsätt et al., 2006) and this is still the only direct evidence of diet that has been reported for any confuciusornithid. However, C. sanctus was probably omnivorous rather than exclusively piscivorous or even faunivorous, since the robust, toothless bill seems capable of cracking seeds (Dalsätt et al., 2006).
C. dui
The specific name of C. dui(Hou et al., 1999) is after a Mr. Du, who collected the holotype specimen and donated it to the ⅣPP for scientific research. Diagnostic features of this species include: smaller than C. sanctus; upper jaw more pointed anteriorly than in C. sanctus; rostral part of mandible slender, without an anteroventral expansion; sternum more elongate than in C. sanctus, with paired short lateral processes and an anterior notch; claw of alular digit not as large as in C. sanctus; intermediate phalanx of manual digit Ⅱ only slightly bowed; tarsometatarsus relatively shorter than in C. sanctus and shorter than pygostyle (Fig. 8).
Figure
8.
Cast of the holotype of Confuciusornis dui, IVPP V11553
C. dui is noteworthy because the holotype was the first described bird of any kind to retain a well-preserved, intact diapsid temporal region, a feature not clearly shown in Archaeopteryx. Given that the bones of the temporal region are fused and reduced in extant birds, the C. dui holotype provided the first unambiguous evidence that a relatively primitive configuration of the temporal region was retained in some basal birds. It is most parsimonious to assume that the same condition is characteristic of Archaeopteryx, even though relatively little direct evidence for this is preserved. Furthermore, C. dui also preserves an impression of a horny rhamphotheca extending anteriorly beyond the tips of the bony upper and lower jaws, representing the most direct available evidence for a horny bill in Confuciusornis.
C. feducciai
The specific name of C. feducciai (Zhang et al., 2009) honors after the American ornithologist Alan Feduccia. The only known specimen displays the following combination of features: larger size than is typical for other species of Confuciusornis; furcula nearly Ⅴ-shaped; sternum broader than long; forelimb to hind limb length ratio about 1.15; proximal end of humerus flattened and thin, lacking fenestra such as present in C. sanctus; first phalanx of alular digit slim; intermediate phalanx of manual digit Ⅱ only slightly bowed; ischium relatively long, about two-thirds length of pubis. However, at least some of these individual features may fall within the range of variation of C. sanctus. In fact, Chiappe et al. (2008) included the specimen that was later used as the holotype of C. feducciai in a morphometric analysis intended to analyze the life history of C. sanctus, this specimen plotted within a growth series established for the latter species.
Changchengornis
The generic name of C. hengdaoziensis(Chiappe et al., 1999; Ji et al., 1999) is from "Chang Cheng", meaning Great Wall in Chinese and the specific name refers to the Hengdaozi Beds, from which the only known specimen was collected. The Hengdaozi Beds lie within what is now regarded as the lower part of the Yixian Formation, although these strata have sometimes been interpreted as belonging to a separate Chaomidianzi Formation (Chiappe et al., 1999). The species differs from other basal bird genera in the following features: strongly curved beak; mandible high caudally and much shorter than skull; deltopectoral crest not perforated by foramen as in C. sanctus; metacarpal Ⅱ twice as long as metacarpal Ⅰ; hallux proportionally longer than in Confuciusornis; caudal margin of sternum forms much steeper "Ⅴ" shape than in C. sanctus; metatarsals Ⅲ and Ⅳ fused distally.
Changchengornis was found by Ji et al. (1999) and Chiappe et al. (1999) to be the sister taxon of Confuciusornis, although it should be noted that the confuciusornithid Jinzhouornis was unknown at the time these analyses were carried out. In the present paper all three derived confuciusornithid genera are considered to form an unresolved trichotomy pending further study. Changchengornis is interpreted to be a partly arboreal and partly terrestrial bird as has been proposed for C. sanctus (Chiappe et al., 1999), despite the presence in Changchengornis of a longer hallux that was presumably better adapted for grasping.
Jinzhouornis
Jinzhouornis is a genus containing the two species J. yixianensis and J. zhangjiyingia (Hou et al., 2002). The generic name refers to Jinzhou City (Fig. 2), within whose boundaries the holotype of J. yixianensis was collected. Jinzhouornis can be identified on the basis of the following combination of features: skull low and long; snout robust and long; orbit moderate in size; part of skull rostral to orbit accounts for more than half of skull length; cervical vertebrae shorter than in Confuciusornis; more than 12 dorsal vertebrae present; manual claws strongly curved; scapula approximately equal in length to humerus. However, Chiappe et al. (2008; see also Chiappe, 2007) stated that the features distinguishing specimens assigned to Jinzhouornis from C. sanctus actually represent variations at the intraspecific level, raising the possibility that both species of Jinzhouornis are junior synonyms of C. sanctus. However, this issue has never been investigated in detail and our observations suggest that each postulated species of Jinzhouornis does possess some diagnostic features. In the present paper we regard both species as provisionally valid pending further study.
J. yixianensis
The specific name of J. yixianensis refers to Yixian County within Jinzhou City (Fig. 2), where the only known specimen came from. It is a medium-sized bird with a long snout, a relatively small braincase, a slender humerus, a proximally fused tarsometatarsus and a slender fifth metatarsal that remains unfused to the rest of the metatarsus, even proximally (Fig. 9).
Figure
9.
Holotype of Jinzhouornis yixianornis, IVPP V14412
The specific name of J. zhangjiyingia also refers to Zhangjiying Township in Beipiao City (Fig. 2), where the specimen came from. It is identifiable by the following features: skull larger than in J. yixianensis; premaxilla extends caudally, contacting frontal; furcula more slender than in C. sanctus; shaft of humerus more robust than in J. yixianensis; lateral margin of proximal end of humerus curved laterally (Fig. 10).
Figure
10.
Holotype of Jinzhouornis zhangjiyinensis, IVPP V 12352
Only fifteen years have elapsed since the first known Chinese basal bird, Confuciusornis, was described (Hou et al., 1995a, 1995b). However, additional discoveries of exquisite specimens, combined with painstaking work by researchers, have made the Chinese fossil record into an astonishing fount of information about the early evolution of birds and flight. Many new species of basal birds have been described in these years and they often show primitive features that highlight their intermediate position in the transition from non-avian theropods to crown-group birds. However, correctly placing the basal birds in their precise phylogenetic positions is sometimes a difficult problem.
Jeholornis typically emerges in phylogenetic analyses as the most primitive Chinese bird, mainly because of its long skeletal tail resembling that of Archaeopteryx. However, one analysis (Zhou et al., 2009) found Sapeornis to be more basal than Jeholornis, implying that the long tail of Jeholornis might have been secondarily re-acquired. Conversely, the coracoid is strut-like in Jeholornis but non-strut-like in Sapeornis and Archaeopteryx and the presence of the primitive coracoid morphology in Sapeornis would be consistent with the more basal phylogenetic position postulated by Zhou et al. (2009).
Although Zhongornis was found by Gao et al. (2008) to be among the most primitive basal birds, this result is uncertain partly because the only known specimen of Zhongornis is a juvenile. It is possible, for example, that the unfused caudal vertebrae would have consolidated into a pygostyle had growth continued, introducing a derived character state not evident in the specimen. This is a common problem in paleontology, albeit one that could be partially solved in this case by the discovery of more basal birds at a range of developmental stages.
Although the published description of Jinzhouornis(Hou et al., 2002) indicates that this taxon is osteologically similar to Confuciusornis and Changchengornis, suggesting a close relationship to these taxa, Jinzhouornis has never been included in a formal phylogenetic analysis. Accordingly, we regard all three confuciusornithid genera as forming a clade whose internal relationships are unresolved, and which is the sister taxon of Eoconfuciusornis (Fig. 1).
It is evident that many questions regarding the structure, evolution and biology of Chinese basal birds remain to be solved. With the discovery of new fossils and the development and application of new technologies, we may expect a more detailed and accurate picture of basal birds and their evolution to emerge in the future.
Acknowledgements
This work was supported by the Ministry of Science and Technology of China, the National Natural Science Foundation of China, and the Chinese Academy of Sciences. We thank two anonymous referees for their constructive comments on our manuscript.
Balbontín, J., De Lope, F., Hermosell, I.G., Mousseau, T.A., Møller, A.P., 2011. Determinants of age-dependent change in a secondary sexual character. J. Evol. Biol. 24, 440-448. .
Buchholz, R., 1996. Thermoregulatory role of the unfeathered head and neck in male wild turkeys. Auk 113, 310-318. .
Carroll, J.M., Davis, C.A., Elmore, R.D., Fuhlendorf, S.D., 2015. A ground-nesting galliform's response to thermal heterogeneity: implications for ground-dwelling birds. PLoS One 10, e0143676. .
Chittenden, H., Allan, D., Weiersbye, I., 2012. Roberts Geographic Variation of Southern African Birds. The John Voelcker Bird Book Fund, Cape Town.
Clancey, P.A., 1980. Checklist of Southern African Birds. The Southern African Ornithological Society, Pretoria.
Cook, T.R., Martin, R., Roberts, J., Häkkinen, H., Botha, P., Meyer, C., et al., 2020. Parenting in a warming world: thermoregulatory responses to heat stress in an endangered seabird. Conserv. Physiol. 8, coz109. .
Cooney, C.R., Seddon, N., Tobias, J.A., 2016. Widespread correlations between climatic niche evolution and species diversification in birds. J. Anim. Ecol. 85, 869-878. .
Cornwallis, C.K., Dean, R., Pizzari, T., 2014. Sex-specific patterns of aging in sexual ornaments and gametes. Am. Nat. 184, E66-E78. .
Crowe, T.M., 1978. The evolution of guinea-fowl (Galliformes, Phasianidae, Numidinae) taxonomy, phylogeny, speciation and biogeography. Ann. S. Afr. Mus. 76, 43-136.
Crowe, T.M., 1979. Adaptive morphological variation in Helmeted Guineafowl Numida meleagris and Crested Guineafowl Guttera pucherani. Ibis 121, 313-320. .
Crowe, T.M., Crowe, A.A., 1979. Anatomy of the vascular system of the heads and neck of Helmeted Guineafowl Numida meleagris. J. Zool. 188, 221-233. .
Crowe, T.M., Elbin, S.B., 1987. Social behaviour of Helmeted Guineafowl Numida meleagris. S. Afr. J. Wildl. Res. 1, 55-57.
Crowe, T.M., Withers, P.C., 1979. Brain temperature regulation in Helmeted Guineafowl. S. Afr. J. Sci. 75, 362-365.
Delhey, K., 2017. Darker where cold and wet: Australian birds follow their own version of Gloger's rule. Ecography 41, 673-683. .
Eduardo, J., Bicudo, P.W., Buttemer, W.A., Chappel, M.A., Pearson, J.T., Bech, C., 2010. Ecological and Environmental Physiology of Birds. Oxford University Press, New York.
Fairbairn, D.J., Blanckenhorn, W.U., Szekely, T., 2008. Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism. Oxford University Press, Oxford. doi: .
Florez-Duquet, M., Mcdonald, R.B., 1998. Cold-induced thermoregulation and biological aging. Physiol. Rev. 78, 339-358.
Fernandez, M.E., Labaque, M.C., Orso, G., Marin, R.H., Kembro, J.M., 2021. Short- and long-term dynamics of the physiological and behavioral response to heat stress and thymol supplementation in Japanese quail. J. Therm. Biol. 97, 102876. .
Friedman, N.R., Miller, E.T., Ball, J.R., Kasuga, H., Remeš, V., Economo, E.P., 2019. Evolution of a multifunctional trait: shared effects of foraging ecology and thermoregulation on beak morphology, with consequences for song evolution. Proc. R. Soc. Lond. B. 286, 20192474. .
Galván, I., Palacios, D., Negro, J.J., 2017. The bare head of the Northern Bald Ibis (Geronticus eremita) fulfills a thermoregulatory function. Front. Zool. 14, 15. .
Galván, I., Rodríguez-Martínez, S., Carrascal, L.M., 2018. Dark pigmentation limits thermal niche position in birds. Funct. Ecol. 32, 1531–1540. .
Gerson, A.R., Smith, E.K., Smit, B., McKechnie, A.E., Wolf, B.O., 2014. The impact of humidity on evaporative cooling in small desert birds exposed to high air temperatures. Physiol. Biochem. Zool. 87, 782-795. .
Grosiak, M., Koteja, P., Bauchinger, U., Sadowska, E.T., 2020. Age-related changes in the thermoregulatory properties in bank voles from a selection experiment. Front. Physiol. 11, 576304. .
Hanan, N.P., Prihodko, L., Ross, C.W., Bucini, G., Tredennick, A.T., 2020. Gridded estimates of woody cover and biomass across Sub-Saharan Africa, 2000-2004. ORNL DAAC, Oak Ridge. .
Hoops, D., Whiting, M.J., Keogh, J.S., 2022. A smaller habenula is associated with increasing intensity of sexual selection. Brain Behav. Evol. .
Iverson, E.N.K., Karubian, J., 2017. The role of bare parts in avian signalling. Auk 134, 587-611. .
Jeren, P., Jenni-Eiermann, S., McKeegan, D., McCafferty, D.J., Nage, R.G.R., 2019. Eye region surface temperature dynamics during acute stress relate to baseline glucocorticoids independently of environmental conditions. Physiol. Behav. 210, 112627. .
Kim, D.H., Lee, Y.K., Lee, S.D., Kim, S.H., Lee, K.W., 2021. Physiological and behavioral responses of laying hens exposed to long-term high temperature. J. Therm. Biol. 99, 103017. .
Kimball, R.T., Edward, L., Braun, J., 2008. A multigene phylogeny of Galliformes supports a single origin of erectile ability in non-feathered facial traits. J. Avian Biol. 39, 438-445. .
Kimball, R.T., St. Mary, C.M., Braun, E.L., 2011. A macroevolutionary perspective on multiple sexual traits in the Phasianidae (Galliformes). Int. J. Evol. Biol. 11, 423938. .
Larochelle, J., Delson, J., Schmidt-Nielsen, K., 1982. Temperature regulation in the Black Vulture. Can. J. Zool. 60, 491-494. .
Little, R., 2016. Terrestrial Gamebirds and Snipes of Africa. Jacana, Cape Town.
Madge, S., McGowan, P., 2002. Pheasants, Partridges and Grouse. Christopher Helm, London.
Møller, A.P., de Lope, F., 1999. Senescence in a short-lived migratory bird: age-dependent morphology, migration, reproduction and parasitism. J. Anim. Ecol. 68, 163-171. .
Moore, A.J., 1990. The evolution of sexual dimorphism by sexual selection: the separate effects of intrasexual selection and intersexual selection. Evolution 44, 315-331.
Mori, E., Mazza, G., Lovari, S., 2017. Sexual dimorphism. In: Vonk, J., Shakelford, T. (Eds. ), Encyclopedia of Animal Cognition and Behavior. Springer International Publishing, Switzerland, pp. 1-7.
.
Nassar, P.N., Jackson, A.C., Carrie, D.R., 2001. Entraining the natural frequencies of running and breathing in guinea fowl (Numida meleagris). J. Exp. Biol. 204, 1641-1651.
Nilsson, J-A., Molokwu, M.N., Olsson, O., 2016. Body temperature regulation in hot environments. PLoS One 11, e0161481. .
Nord, A., Giroud, S., 2020. Lifelong effects of thermal challenges during development in birds and mammals. Front. Physiol. 11, 419. .
Pereyra, M.E., Morton, M.L., 2001. Nestling growth and thermoregulatory development in subalpine Dusky Flycatchers. Auk 118, 116-136. .
Pessato, A., McKechnie, A.E., Buchanan, K.L., Mariette, M.M., 2020. Vocal panting: a novel thermoregulatory mechanism for enhancing heat tolerance in a desert-adapted bird. Sci. Rep. 10, 18914. .
Playà-Montmany, N., González-Medina, E., Cabello-Vergel, J., Parejo, M., Abad-Gómez, J.M., Sanchez-Guzmán, J.M., et al., 2021. The thermoregulatory role of relative bill and leg surface areas in a Mediterranean population of Great tit (Parus major). Ecol. Evol. 11, 15936-15946. .
Prinsloo, H.C., Harley, V., Reilly, B.K., Crowe, T.M., 2005. Sex-related variation in morphology of Helmeted Guineafowl (Numida meleagris) from Riemland of the northeastern Free State, South Africa. S. Afr. J. Wildl. 35, 95-96.
Pyle, P., 2021. Examination of digital images from Macaulay Library to determine avian molt strategies: a case study on molts and plumages in eight species of North American hummingbirds. bioRxiv .
Remeš, V., Martin, T.E., 2002. Environmental influences on the evolution of growth and developmental rates in passerines. Evolution 56, 2505-2518. .
Siegfried, W.R., 1966. Growth, plumage, development and moult in Crowned Guineafowl Numida meleagris coronata Gurney. Depart. Nat. Conserv. Invest. Rep. 9, 1-52.
Stevens, M., Párraga, C.A., Cuthill, I.C., Partridge, J.C., Troscianko, T.S., 2007. Using digital photography to study animal coloration. Biol. J. Linn. Soc. 90, 211-237. .
Stettenheim, P.R., 2000. The integumentary morphology of modern birds-an overview. Am. Zool. 40, 461-477. .
Stuart-Fox, D.M., Ord, T.J., 2004. Sexual selection, natural selection and the evolution of dimorphic coloration and ornamentation in agamid lizards. Proc. R. Soc. Lond. B. 271, 2249-2255. .
Stuart-Fox, D.M., Newton, E., Clusella-Trullas, S., 2017. Thermal consequences of colour and near-infrared reflectance. Phil. Trans. R. Soc. B. 372, 20160345. .
Talbot, W.A., McWhorter, T.J., Gerson, A.R., McKechnie, A.E., Wolf, B.O., 2017. Avian thermoregulation in the heat: evaporative cooling capacity of arid-zone Caprimulgiformes from two continents. J. Exp. Biol. 220, 3488-3498. doi. org/10.1242/jeb. 161653.
Tattersall, G.J., Arnaout, B., Symonds, M.R., 2017. The evolution of the avian bill as a thermoregulatory organ. Biol. Rev. 92, 1630-1656. .
Van Jaarsveld, B., Bennett, N.C., Czenze, Z.J., Kemp, R., van de Ven, T.M., Cunningham, S.J., et al., 2021. How hornbills handle heat: sex-specific thermoregulation in the southern yellow-billed hornbill. J. Exp. Biol. 224, jeb232777. .
Van Niekerk, J.H., 1979. Social and breeding behaviour of the Crowned Guineafowl in the Krugersdorp Game Reserve. Ostrich 50, 188-189.
Van Niekerk, J.H., 1980. Some socio-biological features of Crowned Guineafowl in the Krugersdorp Game Reserve. Bokmakierie 32, 102-108.
Van Niekerk, J.H., 1982. Stalking and enticing guineafowl in a game reserve. Safring 11, 57-61.
Van Niekerk, J.H., 1983. Marking and observing Helmeted Guineafowl in the Krugersdorp Game Reserve. Safring 12, 48-53.
Van Niekerk, J.H., 1985. Observations on courtship in Swainson's Francolin. Bokmakierie 35, 90-92.
Van Niekerk, J.H., 2010. Social organization of a flock of Helmeted Guineafowl (Numida meleagris) at the Krugersdorp Game Reserve, South Africa. Chin. Birds. 1, 22-29. .
Van Niekerk, J.H., 2021. Population ecology of Red-necked Spurfowl Paternities afer in the coastal towns of the Eastern Cape province, South Africa. Ostrich 92, 32–40. .
Van Niekerk, J.H., Barendse, M., Mare, F., 2009. Behaviour of Red-necked Spurfowl Pternistis afer in the Boknes and Cannon Rock coastal resorts, Alexandria district, Eastern Cape province, South Africa. Ostrich 80, 43-47. .
Van Vuuren, A.K.J., Kemp, L.V., McKechnie, A.E., 2020. The beak and unfeathered skin as heat radiators in the Southern Ground-Hornbill. J. Avian Biol. 51, e02457. .
Vignal, A., Boitard, S., Thébault, N., Dayo, G.K., Yapi-Gnaore, V., Youssao Abdou Karim, I., et al., 2019. A guinea fowl genome assembly provides new evidence on evolution following domestication and selection in galliformes. Mol. Ecol. Resour. 19, 997-1014. .
Walsberg, G.E., 1982. Coat color, solar heat gain, and conspicuousness in the phainopepla. Auk 99, 495-502. .
Withers, P.C., Crowe, T.M., 1980. Brain temperature fluctuation in Helmeted Guineafowl under semi-natural conditions. Condor 82, 99-100.
World Bank, 2019. Global Solar Atlas 2.0. Solargis.
Table
1.
List of Chinese basal birds and their geographic and stratigraphic distribution. Holotype specimens are housed at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), Beijing; the Dalian Natural History Museum (D), Dalian, Liaoning Province; and the National Geological Museum of China (GMV), Beijing.


 DownLoad:
DownLoad:














 Email Alerts
Email Alerts RSS Feeds
RSS Feeds